Die komplexe Sinfonie des Sehens der Fruchtfliege
Durch einen neuronalen Mikroschaltkreis kann eine Signalart für mehrere Zwecke genutzt werden
Ein Orchester komplexer neuronaler Netzwerke spielt ein symphonisches Meisterwerk namens Sehen – ein spannendes Forschungsfeld für Neurobiologinnen und Neurobiologen wie Alexander Borst, Direktor am Max-Planck-Institut für biologische Intelligenz. In der jüngsten Studie seiner Abteilung fanden die Forschenden einen Mikroschaltkreis, der aktivierende in hemmende Signale umwandelt und so eine einzige Art von neuronalem Input für mehrere Zwecke nutzen kann. Die Entdeckung dieses Mikroschaltkreises ist ein wichtiges Puzzlestück zum besseren Verständnis des Sehprozesses der Fruchtfliege Drosophila und letztlich des Sehens an sich.

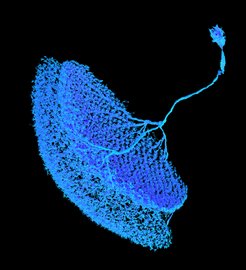
Das Sehen ist einer der wichtigsten Sinne von uns Menschen. Entsprechend groß ist der Anteil des Gehirns, der dem Verarbeiten visueller Informationen gewidmet ist. Um visuelle Informationen schnell und akkurat zu verarbeiten, muss eine Vielzahl neuronaler Netzwerke ein komplexes Zusammenspiel leisten. Beim Verständnis dieses Zusammenspiels helfen Fruchtfliegen. Denn erstaunlicherweise gibt es neben den offensichtlichen Unterschieden zwischen den Augen von Fliegen und Menschen auch viele Parallelen in der Art und Weise, wie die Gehirne visuelle Informationen verarbeiten. Da das visuelle System der Fliegen sehr effizient, aber deutlich weniger komplex als das unsere ist, überrascht es nicht, dass dies eines der am besten verstandenen Nervenzellnetzwerke in den Neurowissenschaften ist.
Im visuellen System von Fruchtfliegen wandelt eine Kaskade von Zellen Lichtinformationen in richtungsspezifische Signale um. T4-Zellen zum Beispiel reagieren auf helle, sich bewegende Kanten (ON-Pfad), während T5-Zellen auf sich bewegende dunkle Kanten reagieren (OFF-Pfad). Sowohl die T4- als auch T5-Zellen haben vier Untertypen, die jeweils auf eine der vier Kardinalrichtungen (von hinten nach vorne, von vorne nach hinten, von oben nach unten, sowie von unten nach oben) abgestimmt sind. Das bedeutet, dass jede Nervenzelle nur auf visuelle Reize aus einer ganz bestimmten Richtung reagiert, die sogenannte Vorzugsrichtung. Auf Bewegungen in eine andere Richtung (Nullrichtung) zeigt sie keine Reaktion.
Erst kürzlich hatte die Abteilung den Mechanismus zur Berechnung der Richtungsselektivität im ON-Pfad entschlüsselt. Die genaue Umsetzung im OFF-Pfad blieb jedoch bisher noch unerforscht. Um dies zu ändern haben die Wissenschaftler*innen der Abteilung von Alexander Borst nun eine einzigartige Kombination von Kalzium-Bildgebung mit thermogenetischen, optogenetischen und pharmakologischen Techniken entwickelt, um das neuronale Netzwerk des OFF-Pfades in Drosophila zu untersuchen.
Das Team fand heraus, dass mehrere Eingangsneuronen die T5-Zellen aktivieren, sobald eine Bewegung entlang der bevorzugten Richtung der T5-Zellen wahrgenommen wird. Daraufhin testeten die Forschenden die Rolle dieser Eingangsneuronen für die Richtungsberechnung der T5-Zellen, indem sie die Eingangsneuronen entweder unterdrückten oder aktivierten. Die Ergebnisse zeigten, dass die Eingangsneuronen für die Feinabstimmung der Vorzugsrichtung der T5-Zellen verantwortlich sind.
Neben den aktivierenden Eingangsneuronen gibt es auch hemmende Neuronen, die Signale reduzieren. Auf diese Weise werden in den T4-Zellen alle Signale aus anderen Richtungen als der Vorzugsrichtung unterdrückt. Überraschenderweise gibt es für T5-Zellen keine solchen hemmenden Neuronen. Dies stellte die Forschenden vor ein Rätsel: Wie funktioniert die Unterdrückung der Nullrichtung in T5? Mit anderen Worten: Wie können diese Nervenzellen ganz ohne hemmende Neuronen nur auf Signale ihrer bevorzugten Richtung reagieren?
Das Team, geleitet von Amalia Braun, entdeckte einen Mikroschaltkreis, welcher der Unterdrückung der Nullrichtung in T5-Zellen zugrunde liegt: CT1, ein ungewöhnliches Großfeldneuron, empfängt aktivierende Signale und wandelt diese in hemmende Signale um, die wiederum die Unterdrückung der Nullrichtung ermöglichen. “Mit dem Einsatz genetischer Methoden konnten wir einzelne Zelltypen entfernen und so zeigen, dass sich die Abstimmungskurve der Richtungserkennung einer T5-Zelle erheblich verbreitert, wenn CT1 wegfällt. Dies konnte auch mit Hilfe algorithmischer Modellierung unterstrichen werden", erklärt Amalia Braun. "Während also die aktivierenden Neuronen die T5-Zellen direkt für ihre bevorzugte Richtung beeinflussen, führt ein Umweg über eine CT1-Zelle zur Umkehrung des Signals und so zur Nullrichtungsunterdrückung.”
Mit der Entdeckung dieses Mikroschaltkreises konnte die Abteilung der komplexen Komposition des visuellen Systems von Drosophila einen weiteren Satz hinzufügen. "Der CT1-Mikroschaltkreis ist ein eindrucksvolles Beispiel für die vielschichtigen und unterschiedlichen Mechanismen der Hemmung und Erregung im Gehirn und dafür, wie ein neuronales Netzwerk einen einzigen Signaltyp für mehrere Zwecke nutzen kann", fasst der Letztautor der Studie, Matthias Meier, zusammen.
Die Identifizierung dieses Mikroschaltkreises, der die Bewegungserkennung ermöglicht und prägt, vertieft erheblich unser Verständnis der visuellen Verarbeitung von Drosophila – und letztlich des Sehens an sich.