Forschungsbericht 2015 - Max-Planck-Institut für biologische Intelligenz (Standort Martinsried)
Der Schaltplan des Gehirns
Das einzige Ziel des Denkens ist es letztlich zu berechnen, welche von allen möglichen Handlungen dem Organismus am meisten nützt und wie diese Handlung am besten in eine Abfolge von Muskelkontraktionen umzusetzen ist. Im Zentrum aller Rechenvorgänge steht der Fluss von Informationen. Dieser hängt wiederum davon ab, wie die einzelnen Schaltelemente – im Gehirn die Nervenzellen – miteinander verbunden sind. Wenn wir wissen, "wer mit wem in Verbindung steht", können wir der Information im Gehirn folgen und bestimmen, welche Signale miteinander wechselwirken können und welche nicht. Weil die Zellen und ihre Verbindungen den Informationsfluss nicht nur ermöglichen, sondern auch im Einzelnen kontrollieren, stellen sie nicht nur die physikalische Grundlage für das Rechnen des Gehirns dar. Sie enthalten auch die Rechenalgorithmen, also die "Software" des Gehirns.
Im Unterschied zu den meisten anderen Zellen eines Organismus, von denen jede nur etwa ein Dutzend Nachbarn hat, berührt jede Nervenzelle tausende – in Extremfällen sogar hunderttausende – anderer Nervenzellen. Mit vielen dieser Zellen bestehen sogenannte synaptische Verbindungen, über die eine Zelle Signale empfängt oder aussendet. Synapsen können sich chemischer Botenstoffe bedienen oder direkte elektrische Verbindungen herstellen und sind der Weg, auf dem Zellen miteinander kommunizieren – das heißt, sich gegenseitig in ihrer Aktivität beeinflussen. Um eine so große Zahl von anderen Zellen berühren zu können, wachsen aus jeder Nervenzelle mehrere schlanke, oft vielfach baumartig verzweigte Fortsätze heraus. Diese werden, je nachdem, ob sie Signale senden oder empfangen, Axone und Dendriten genannt und sind die "Drähte" des Gehirns. Fortsätze vieler Zellen bilden dann ein verschlungenes, durch die Synapsen in sich verbundenes Netzwerk – das "Neuropil". Schon der spanische Neurowissenschaftler Santiago Ramon Y Cajal (1852-1934), einer der Begründer der klassischen neuroanatomischen Forschung, hat vieles über die Form der Nervenzellen und manches über ihre Verbindungen herausgefunden. Da es damals aber nur Lichtmikroskope gab, mit denen die Synapsen und Drähte im dicht gepackten Neuropil nicht aufgelöst werden können, musste er sich eines Anfärbetricks des Italieners Camillo Gogi (1843-1926) bedienen, der einzelne Zellen zusammen mit all ihren Drähten sichtbar macht. So konnte Cajal sich zwar ein sehr gutes Bild des Verzweigungsmusters einzelner Zelle machen, aber für Aussagen über Verbindungen waren er und andere Neuroanatomen vielfach auf Vermutungen angewiesen.
Konnektomik – das Verbindungsmuster bestimmen
Mit der viel besseren, im Nanometerbereich (10-9 m) liegenden Auflösung der in den 1940er Jahren erfundenen Elektronenmikroskopie (EM) ist es nun möglich, jede Synapse zu erkennen und jedem Draht zu folgen und damit im Prinzip das komplette Verbindungsmuster (Konnektom) eines Gehirns zu bestimmen. Das Elektronenmikroskop wurde in der Tat auch benutzt, um das Verbindungsdiagramm eines kompletten Nervensystems zu bestimmen: das des Fadenwurms Caenorhabditis elegans [1]. Obwohl das "Gehirn" von C. elegans nur ein paar hundert Nervenzellen und ein paar tausend Synapsen hat, waren die Wissenschaftler damit über zehn Jahre lang beschäftigt. Ein Mäusegehirn hat dagegen etwa hundert Millionen (108) Zellen, die durch etwa 100 Milliarden (1011) Synapsen miteinander verbunden sind. Im menschlichen Gehirn sind es sogar 100 Milliarden Zellen, verbunden mit einer Billiarde (1015) Synapsen.
Ist damit die Bestimmung des Konnektoms komplexerer Nervensysteme von vornherein aussichtslos? Nicht notwendigerweise, wie die Entwicklung der Gensequenzierung gezeigt hat. Allein in den letzten 15 Jahren sind die Kosten pro Genom durch die Erfindung neuer und der Verfeinerung bestehender Methoden um einen Faktor 100 000 gefallen. Mithilfe neuer Methoden [2] – manche davon in dieser Abteilung entwickelt [3] – wurden im Laufe des letzten Jahrzehnts die Verbindungsdiagramme kleinerer Bereiche komplexerer Nervensysteme erstellt. Sie haben zum Beispiel zum Verständnis des Bewegungssehens in der Netzhaut beigetragen [4; 5]. Bevor jedoch der Schaltplan eines ganzen Gehirns (selbst eines so kleinen wie das der Maus) erstellt werden kann, müssen eine ganze Reihe von Hindernissen überwunden werden. Das erste davon ist die Herstellung eines geeigneten Ganzhirn-Präparates.
Die Färbung eines ganzen Gehirns


Abb.1: Schnitt in vertikaler und rechts-links-Richtung durch Mäusehirn, das mithilfe der BROPA-Methode vorbereitet und mit einem Rasterelektronenmikroskop abgebildet wurde. Der Kalibrierungsbalken ist einen Millimeter lang.
In einem für die Konnektomik geeigneten Gehirnpräparat müssen die zellulären, besonders die synaptischen Feinstrukturen überall erhalten sein. Auch müssen die Färbung mit Schwermetallen und die Einbettung in Kunstharz gleichmäßig sein. Das erste ist entscheidend für die Erhaltung aller Verbindungsdaten aus dem lebenden Tier. Das zweite, die gleichmäßige Färbung, sorgt für einen ausreichenden Kontrast bei der Abbildung im Elektronenmikroskop. Die Einbettung erlaubt das zuverlässige Abschneiden dünner Scheibchen, wie es für die Auflösung der dritten Raumdimension notwendig ist. Mit einer Kombination von Geduld und sorgfältiger Erkundung verschiedener chemischer Verbindungen und Reaktionsverfahren ist es gelungen, die sog. BROPA-Methode (brain-wide reduced-osmium staining with pyrogallol-mediated amplification method) zu entwickeln, die all diese Bedingungen erfüllt [6], (Abb. 1). Zusätzlich zu einer Verlängerung der Verweilzeiten in den verschiedenen Lösungen, die den Zu- und Abtransport verschiedener Substanzen an allen Punkten der Probe allein durch Diffusion erlaubt, musste jedoch an einigen Stellen auch die Lösungszusammensetzung geändert werden. So war es zum Beispiel nötig, einen Ersatz für Thiocarbohydrazid zu finden, welches bei der Kontrastverstärkung eingesetzt wurde, aber gleichzeitig das Gewebe durch die Bildung winziger Gasbläschen beschädigte. Der Ersatzstoff, Pyrogallol, wurde durch Serienexperimente aus einem Katalog von Substanzen ausgewählt. Ein weiteres Problem, die Entstehung einer oberflächennahen Diffusionsbarriere, konnte durch die Beifügung von Formamid, einem Derivat der Ameisensäure, behoben werden [6].
 Rekonstruktion eines mit synaptischen Dornen besetzten Dendritenendstücks (grün) aus dem \"Striatum\", zusammen mit 74 ankommenden synaptischen Verbindungen. Das Striatum ist im Gehirn für die Verschaltung gezielter Bewegungen zuständig. (b) Querschitte durch eine erregende Synapse, einmal in der Abbildungsebene (xy) und einmal senkrecht dazu (yz) aus einem mittels der serial blockface electron microscopy aufgenommenen Bilderstapel (Auflösung 10 x10 x30 nm) durch ein Teilstück eines nach der BROPA-Methode vorbereiteten Gehirns. (c) xy-Schnitt durch eine hemmende Synapse.")
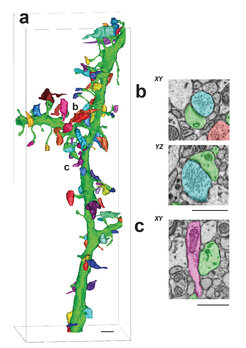
Abb. 2: (a) Rekonstruktion eines mit synaptischen Dornen besetzten Dendritenendstücks (grün) aus dem "Striatum", zusammen mit 74 ankommenden synaptischen Verbindungen. Das Striatum ist im Gehirn für die Verschaltung gezielter Bewegungen zuständig. (b) Querschitte durch eine erregende Synapse, einmal in der Abbildungsebene (xy) und einmal senkrecht dazu (yz) aus einem mittels der serial blockface electron microscopy aufgenommenen Bilderstapel (Auflösung 10 x10 x30 nm) durch ein Teilstück eines nach der BROPA-Methode vorbereiteten Gehirns. (c) xy-Schnitt durch eine hemmende Synapse.
Obwohl das Gewebe dem Augenschein nach gut erhalten war, wurde in Testreihen sichergestellt, dass mit der BROPA-Methode die Strukturinformation, die zum Verfolgen der Drähte und der Identifizierung der Synapsen nötig ist, erhalten bleibt. Dazu verfolgten Wissenschaftler einzelne Axone und Dendriten in einer Reihe von im Gehirn verteilter Testvolumina (Abb. 2). Derzeit prüfen die Martinsrieder Wissenschaftler in Zusammenarbeit mit mehreren Forschungsgruppen in den USA und Japan, ob die mit dem BROPA-Verfahren hergestellten Proben auch für andere, auf der Elektronenmikroskopie basierende Verfahren zur neuronalen Schaltplananalyse geeignet sind.
Schneiden, Abbilden und Analysieren
Die verbleibenden Hindernisse auf dem Weg zu einem kompletten Ganzhirndatensatz sind das Testen und "Debugging" eines speziell zu diesem Zweck entworfenen und in der feinmechanischen Werkstatt des Max-Planck-Instituts gefertigten Ultramikrotoms und seiner Verbindungskammer mit einem Mehrfachstrahl-Elektronenmikroskop, entwickelt von dem bekannten Mikroskophersteller Carl Zeiss. Auch nachdem die Hindernisse auf dem Weg zur Gewinnung eines EM-basierten Volumendatensatzes überwunden sind, wird es eine erhebliche Herausforderung sein, aus diesem Datensatz mit seinen bis zu hundert Billiarden Datenpunkten Erkenntnisse über das Denken zu gewinnen. Schon die Verfolgung der neuronalen Drähte, die für kleinere Volumina noch manuell, wenngleich erheblich vereinfacht durch computergestützte Visualisierungsverfahren erfolgt ist [7], (http://knossostool.org), wird im Gesamtgehirn nur durch eine weitgehende Automatisierung möglich sein. Dies wird nicht nur die Entwicklung neuer Bilderkennungsalgorithmen erfordern, sondern auch den Einsatz von Rechnern in einem Umfang nötig machen, der den für die Analyse des Genoms benötigten noch weit übertreffen könnte. Man kann wohl davon ausgehen, dass so ein Datensatz es ermöglichen wird, nicht nur neue Ideen zu einwickeln, sondern auch viele falsche Theorien auszusondern – also all jene, welche ein Verdrahtungsmuster voraussetzen, das mit dem vorgefundenen nicht übereinstimmt.
Literaturhinweise
Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 314(1165), 1-340 (1986)
Plos Biology 2(11), 1900-1909 (2004)
Nature 500(7461), 168-174 (2013)
Nature Methods 12(6), 541-546 (2015)
Nature Neuroscience 14(8), 1081-U1189 (2011)