Forschungsbericht 2009 - Max-Planck-Institut für medizinische Forschung
Dynamik von Acetylcholinrezeptoren in neuromuskulären Synapsen in vivo
Gruppe Witzemann / Koenen - Molekulare Anatomie neuromuskulärer Synapsen (Prof. Dr. Veit Witzemann)
MPI für medizinische Forschung, Heidelberg
. (A) Motorneuronen (grüne Fluoreszenz durch Färbung nervenspezifischer Neurofilamente) bilden spezifische Kontakte mit Muskelfasern (blaue Fluoreszenz markiert Muskel-spezifische Myosine). An den Kontaktstellen in den Muskelfasern bilden sich postsynaptische Spezialisierungen, die motorischen Endplatten (rote Fluoreszenz zeigt eine Endplatte durch Markierung der postsynaptischen AChR mit Rhodamin-Bungarotoxin). B) Schematische Darstellung des AChR-Komplexes bestehend aus zwei α- und je einer β-, δ-, und γ- (embryonal) oder ε- (adult) Untereinheit. Alle Untereinheiten enthalten vier Transmembranbereiche M1-M4, wobei die M2-Bereiche den Ionenkanal bilden (Skalierung: 20 μm).")

Synapsen sind spezialisierte Kontakte zwischen zwei Nerven oder einem Nerv und einer nicht neuronalen Zelle, wie zum Beispiel die neuromuskuläre Synapse (NMS) in Skelettmuskeln. Mithilfe der Synapsen werden komplexe neuronale Netzwerke angelegt, durch die Lernen, Gedächtnis und insgesamt die Kontrolle aller Lebensfunktionen, einschließlich der Bewegung und Atmung, ermöglicht und gesteuert werden.
Die NMS (Abb. 1A) hat als lange bekanntes Modellsystem entscheidend zur Aufklärung der chemischen Neurotransmission durch Acetylcholin beigetragen. Der nikotinische Acetylcholinrezeptor (AChR) war dann auch der erste Neurotransmitterrezeptor und Ionenkanal, der strukturell und funktionell charakterisiert werden konnte, und der erste Rezeptor, der mit gentechnischen Methoden untersucht wurde.
Rezeptor-induzierte postsynaptische Ströme werden entwicklungsabhängig schneller
 Schematische Darstellung der γ-GFP-Untereinheit mit GFP im cytoplasmatischen Loop. B) Die modifizierte Untereinheit wird exprimiert und in den AChRγ-GFP-Komplex eingebaut, der dann korrekt zur postsynaptischen Membran transportiert wird. Endplatten im Diaphragmamuskel sind während der frühen postnatalen Entwicklung direkt anhand der Grünfluoreszenz sichtbar. C) Endplatten wie in B, dargestellt durch Rhodamin-Bungarotoxin markierte AChR. (Skalierung 20 μm)")

Im Muskel enthalten die embryonalen AChR (AChRγ) zwei α-, und je eine β-, γ- und δ- Untereinheit. Die γ-Untereinheit wird in adulten Rezeptoren (AChRε) durch die ε-Untereinheit ersetzt (Abb. 1B) [1]. Die Kanalkonversion hat zur Folge, dass die "Kanaloffenzeit" verkürzt und die Ionenleitfähigkeit erhöht wird und eine schnellere Signalübertragung zur Optimierung der Feinmotorik ermöglicht wird. Darüber hinaus haben die beiden Rezeptortypen noch weitere unterschiedliche Funktionen: AChRε werden weitgehend Synapsen-spezifisch exprimiert und garantieren die hohe synaptische Konzentration und Stabilität der postsynaptischen Struktur [2]. Die AChRγ haben andererseits die Aufgabe, synaptische Kontaktstellen während der Entwicklung an der richtigen Stelle auf der Muskelfaser zu lokalisieren [3]. Somit sind die Rezeptoren nicht nur Signalvermittler, sondern haben zusätzliche regulatorische Funktionen bei der Bildung und Stabilisierung von Synapsen.
Ein Ziel war es, die AChR-Kanalkonversion direkt zu verfolgen, um zu sehen, wie neue Rezeptoren in die postsynaptische Membran eingebaut werden. Dazu wurde eine gentechnisch veränderte Maus hergestellt, die AChR γ-GFP/γ-GFP-Maus [4], in der das grün fluoreszierende Protein (GFP) in das Gen der AChR γ-Untereinheit eingefügt wurde (Abb. 2A).
Muskel- versus Nerv-induzierte Kanalkonversion
Beobachtungen, dass ε-enthaltende Rezeptoren in Mäusen erst nach der Geburt in signifikanter Zahl auftreten, streng auf postsynaptische Bereiche begrenzt bleiben und ihre Expression durch Faktoren neuronalen Ursprungs experimentell induziert werden kann, sprachen für eine neuronal induzierte Kanalkonversion. Dass auch muskelspezifische Faktoren die Expression von AChRε induzieren, stellt den Einfluss von neuronalen Faktoren in Frage. In AChR γ-GFP/γ-GFP-Mäusen konnte der Austausch der AChRγ/AChRε-Proteinkomplexe direkt in vivo verfolgt werden. Wegen einer stark reduzierten γ-GFP-Expression und unveränderter Expression der ε-Untereinheit findet die Kanalkonversion zu einem früheren Zeitpunkt statt als in Wildtyp-Mäusen. Das heißt, dass allein die Verfügbarkeit der entsprechenden Untereinheiten den Rezeptoraustausch bestimmt, der unabhängig von neuronalen Signalen durch muskelspezifische Mechanismen gesteuert wird [4] .
Direkte Visualisierung der AChR-Kanalkonversion
")
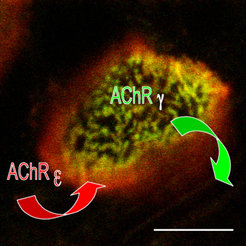
In frühen postnatalen Stadien kann man die Abnahme der grün-fluoreszierenden AChRγ-GFP und die Zunahme der Rhodamin-Bungarotoxin markierten, rot-fluoreszierenden AChRε in den neuromuskulären Synapsen direkt verfolgen. Die neuen Rezeptoren werden in die peripheren Endplattenbereiche integriert und dann zielgerichtet weiter ins Zentrum transportiert. Dieser Austausch erfolgt nicht gleichmäßig über den ganzen Muskel in allen Endplatten, sondern individuell, Muskelfaser-spezifisch in den einzelnen Endplatten.
Rezeptorstabilität ist abhängig von Muskelaktivität
Es ist seit langer Zeit bekannt, dass die metabolische Stabilität der Rezeptoren mit der Entwicklung der NMS zunimmt und die Halbwertszeit t½ von 1 Tag auf 10 Tage steigt. Damit wird scheinbar ein Verlust an Plastizität in Kauf genommen. Neuere Erkenntnisse haben gezeigt, dass die Stabilisierung nicht statisch ist, sondern auf einem aktivitätsabhängig regulierten AChR recycling-Prozess basiert [5].
Einen experimentellen Ansatz zur Analyse bieten Denervierungsexperimente, in denen die Motorneuronen operativ durchtrennt werden. Die Inaktivierung der Muskeln führt zu einem stark erhöhten metabolischen Umsatz der Rezeptoren. Vor allem die embryonale γ-Untereinheit, die im innervierten adulten Muskel reprimiert ist, wird nun transient wieder sehr stark exprimiert, sodass embryonale Rezeptoren die adulten Rezeptoren in den Endplatten ersetzen. Das legte den Schluss nahe, dass die AChR-Stabilität durch unterschiedlich stabile AChR-Typen bestimmt wird. Paradox erschien die Beobachtung, dass Rezeptordichte und Endplattenstruktur nach Denervierung 10-20 Tage unverändert stabil bleiben sollten, obwohl synaptische Rezeptoren schon 3 Tage nach Denervierung schnelle Umsatzraten zeigten.
Mit den AChR γ-GFP/γ-GFP-Mäusen kann man die Denervierungs-induzierten Veränderungen direkt verfolgen. Wie bei der postnatalen Kanalkonversion sieht man, dass neue Rezeptoren gerichtet in die Synapse eingebaut werden (Abb. 4).
 enthalten vorwiegend AChRγ-GFP, die die AChRε ersetzt haben. Die neuen AChRγ-GFP werden im subsynaptischen sarkoplasmatischen Retikulum zusammengebaut (grüne Pfeile) (Skalierung 20 μm).")

Die erhöhte Expression von AChRγ-GFP ist ungefähr fünfzehn Tage nach der Denervierung wieder stark abgefallen, und alle AChRγ-GFP sind wieder durch AChRε ersetzt worden. Diese Ergebnisse zeigen, dass der Einbau/Abbau der Rezeptoren und die synaptische Stabilität der Rezeptoren nicht über unterschiedliche Rezeptortypen reguliert oder durch putative neuronale Komponenten beeinflusst wird, sondern in erster Linie vom Angebot an neu synthesisierten Rezeptoren abhängt. Das heißt, Muskeln stellen ein Gleichgwicht (Synthese/synaptische Stabilität/Abbau) her, das die AChR-Dichte an Synapsen regelt. Dieses Gleichgewicht ist muskelspezifisch und hängt von Aktivitätszustand, bzw. Innervierungszustand ab.
Inzwischen wurde eine neue In-vivo-Analysemethode entwickelt, mit der es möglich ist, Prozesse wie die Kanalkonversion und Veränderungen der metabolischen Stabilität im Muskel über bis zu 8 Stunden direkt zu verfolgen. Damit kann die Dynamik der Acetylcholinrezeptoren in den neuromuskulären Synapsen mit einer bisher nicht erreichten Auflösung untersucht werden.