Forschungsbericht 2010 - Max-Planck-Institut für molekulare Physiologie
Kräftemessen in der Nervenzelle – Wie entwickelt sich die komplexe Form von Neuronen?
Max-Planck-Institut für molekulare Physiologie, Dortmund
Einleitung
Die Form von tierischen Zellen wird primär durch faserartige Proteinstrukturen geprägt, welche eine Art Skelett bilden. Im Gegensatz zu dem starren Skelett von Tier und Mensch ist das Skelett in Zellen – das sogenannte Zytoskelett – nicht nur viel flexibler, sondern auch sehr dynamisch. Insbesondere zwei Arten des Zytoskeletts, die sogenannten Mikrotubuli und die Aktinfilamente, sind aus einzelnen, miteinander verbundenen Einheiten aufgebaut, welche durch Polymerisation und Depolymerisation in einer Zeitspanne von Minuten umgebaut werden können. Diese Dynamik ermöglicht es den Zellen, flexibel auf äußere Stimuli durch Zellformänderungen zu reagieren. Formänderungen von Zellen sind wiederum die Basis für makroskopische Prozesse, beispielsweise in der Embryonalentwicklung. Die Mechanismen, welche diesen dynamischen Auf- und Abbau steuern, sind noch nicht genau verstanden und Gegenstand aktueller Forschung – insbesondere weil deren Störung zu Krebs oder entwicklungsbedingten Krankheiten führen kann.
 und Aktinfilamente (rot), wurden in Neuronen vor (links) und nach (rechts) der Bildung von Neuriten (Zellfortsätzen) angefärbt. Unten: Vergrößerter Bereich eines Wachstumskegels, in welchem Mikrotubuli und Aktin interagieren (Farbkombination im überlagerten Bild von Rot+Grün=Gelb).")
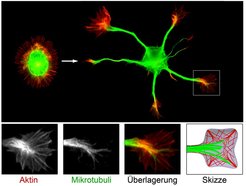
Die Gehirnentwicklung stellt aufgrund ihrer Komplexität einen besonders hohen Anspruch an die präzise räumlich-zeitliche Koordination von Zellwachstum, Migration und Formänderungen. Selbst kleine Veränderungen in den zugrundeliegenden Kontrollmechanismen können zu schweren Krankheiten wie z. B. dem fetalen Alkoholsyndrom, Autismus oder Schizophrenie führen. In Abbildung 1 sind die beiden Hauptkomponenten des Zytoskeletts, die Mikrotubuli und Aktinfilamente, in zwei frühen Entwicklungsstadien von Nervenzellen angefärbt. Zunächst sind die Aktin- und Mikrotubulifilamente relativ symmetrisch angeordnet. Hieraus bilden sich mehrere Zellfortsätze, die sogenannten Neuriten, welche einen mikrotubulibasierten Stamm und ein aktinreiches Ende, den sogenannten Wachstumskegel, aufweisen [1]. Obwohl die Mikrotubuli hauptsächlich im Zellinneren und die Aktinfilamente in der Zellperipherie lokalisiert sind, überlappen diese Strukturen in einem Übergangsbereich (Abb. 1).
Aktinfilamente verschieben Mikrotubuli zum Zentrum der Zelle
Statische Bilder können nur einen sehr eingeschränkten Einblick in das komplexe Verhalten des dynamischen Zytoskeletts bieten. Es ist jedoch möglich, Komponenten des Zytoskeletts in lebenden Zellen durch gentechnisch vermittelte Fusion mit fluoreszierenden Proteinen zu markieren (Abb. 2). Durch anschließende videomikroskopische Untersuchungen kann dann die dynamische Reorganisation dieser Komponenten beobachtet werden.
 werden von rückwärtsgerichteten Kräften aus dem Aktinfluss (türkisfarbene Pfeile) zum Zellzentrum geschoben. Eine größere Anzahl von Mikrotubuli (weiße Pfeile) kann die Position des Aktinflusses modulieren.")
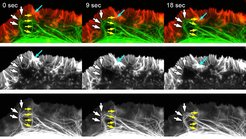
Hier ist zunächst ein ständiger Fluss von Aktinfilamenten vom Zellrand zum Zellzentrum auffällig, der sogenannte retrograde (rückwärtsgerichtete) Fluss (Abb. 2, türkisfarbene Pfeile) [2]. Dieser Fluss entsteht durch die Polymerisation von Aktin am äußersten Zellrand und wird durch die Verschiebung von Aktin durch den molekularen Motor Myosin verstärkt [3]. Der retrograde Aktinfluss kann auch Mikrotubuli in Richtung des Zellinneren verschieben, wenn diese in die aktinreiche äußere Zellregion eintreten [1, 4]. In Abbildung 2 ist dies anhand einer Formänderung der Mikrotubuli erkennbar: Die Kraft, welche über den retrograden Fluss übertragen wird, führt zur Kompression der Mikrotubuli und lässt diese einknicken (Abb. 2, gelbe Pfeile). Dies zeigt, dass Aktinfilamente die Mikrotubuliorganisation direkt beeinflussen können.
Umgekehrt können aber auch Mikrotubuli die Aktinfilamente beeinflussen [1]. In Abbildung 2 begrenzt beispielsweise eine Ansammlung von mehreren Mikrotubuli (weiße Pfeile) die räumliche Anordnung der Aktinfilamente. Es ist jedoch unklar, welche Eigenschaft der Mikrotubuli diesen Einfluss auf das Aktinzytoskelett ausübt. Beispielsweise können Mikrotubuli lokale chemische Signale produzieren, welche die Dynamik von Aktinfilamenten beeinflussen. Alternativ könnte die mechanische Stabilität der Mikrotubuli den Fluss von Aktinfilamenten durch einen eher passiven Mechanismus begrenzen [5]. Mikrotubuli könnten jedoch auch eine aktivere Rolle spielen und durch molekulare Motorproteine gegen den retrograden Fluss geschoben werden. Diese konkurrierenden Kräfte würden zu einer Art Tauziehen führen. Solange keine der Kräfte überwiegt, sind Änderungen in der Organisation des Zytoskeletts und der Zellform minimal.
Molekulare Motoren verschieben Mikrotubuli: Räumliche Organisation und Fokussierung von Kräften durch Selbstorganisation
Durch Zugabe von pharmakologisch aktiven Substanzen kann jedoch von außen künstlich ein Zustand fern eines solchen Kräftegleichgewichts hergestellt werden. In einem solchen Zustand kann der Einfluss von Kräften direkt beobachtet werden. Zum Beispiel kann ein eher unbewegliches Netzwerk von zellulären Mikrotubuli durch Depolymerisation und nachfolgende Repolymerisation in eine Ansammlung von kleinen, frei beweglichen Mikrotubulifragmenten umgewandelt werden. Durch die freie Beweglichkeit der Mikrotubulifragmente können diese leicht durch eine einwirkende Kraft in der Zelle verschoben werden. Tatsächlich haben wir beobachtet, dass eine ursprünglich zufällig verteilte Anordnung von Mikrotubulifragmenten innerhalb von wenigen Minuten reorganisiert wird und nach außen gerichtete Ansammlungen bildet, welche wiederum kleine Zellauswüchse induzieren (Abb. 3) [6].
Diese Reorganisation ist abhängig von einem molekularen Motor, dem sogenannten zytoplasmatischen Dynein. Dieser Motor spielt eine wichtige Rolle in dem gerichteten Transport von zellulärem Transportgut. Er erkennt dabei eine Asymmetrie in der Mikrotubulistruktur, an welcher er sich orientieren kann. Hier sind jedoch Mikrotubuli selbst das Transportgut, welches von Dynein direktional in der Zelle verschoben wird. Dies ist möglich, weil ein Teil der Dynein-Motoren an einer Gerüststruktur in der Plasmamembran verankert ist und daher frei bewegliche Mikrotubuli gegen diese stabile Struktur verschieben kann [6].

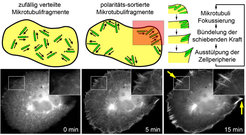
Diese direktionale Verschiebung findet so lange statt, bis alle Mikrotubuli auf ein unüberwindbares Hindernis stoßen. Dies ist meist der Zellrand, an dem sich mehrere Mikrotubulifragmente ansammeln und durch ihre gebündelte Kraft eine kleine Membranausbuchtung induzieren (Abb. 3). Interessanterweise kann sich diese Bildung der Ausbuchtung selbst verstärken: Je mehr Mikrotubulifragmente auf eine Position der Zellperipherie fokussiert sind, desto größer wird die Ausbuchtung und desto mehr der ursprünglich zufällig verteilten Mikrotubulifragmente werden in dieser vergrößerten Ausbuchtung durch seitliche Verschiebung eingefangen. Es handelt sich also um einen Prozess, in welchem die entstehende Struktur – die fokussierten Mikrotubuli – ein integraler Bestandteil eines selbst verstärkenden Mechanismus ist. Dieser Prozess hat Ähnlichkeit zu dem Entstehungsprinzip von Termitenbauten [7, 8]. Die grundlegenden Strukturen dieser makroskopischen Bauten – Säulen und Tunnel aus Lehm – sind ebenfalls ein integraler Bestandteil eines selbst verstärkenden Mechanismus, in welchem Termiten auf der Basis der lokalen Konzentration eines Botenstoffs (Pheromons) in dem sich entwickelnden Bauwerk agieren.
Zusammengenommen kann der vom zytoplasmatischen Dynein vermittelte direktionale Transport eine zunächst zufällig angeordnete Ansammlung von Mikrotubulifragmenten entsprechend ihrer Polarität sortieren und an der Zellperipherie fokussieren. Diese fokussierten Mikrotubulifragmente produzieren eine Kraft, die vom Zellinneren zur Zellperipherie gerichtet ist.
Entgegengesetzte Kräfte von Mikrotubuli und Aktin führen zu einem Tauziehen in Zellen: Implikationen für Zellformänderungen
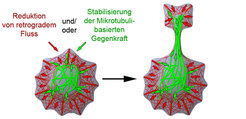
Das oben beschriebene Experiment stellt ein vereinfachtes zelluläres Modellsystem dar, in dem eine auf das Zytoskelett wirkende Kraft deutlich gemacht wird. In sich entwickelnden Neuronen steht diese Kraft, welche auf Mikrotubuli wirkt, jedoch im Gleichgewicht mit einer entgegengesetzten Kraft aus dem retrograden Fluss von Aktinfilamenten. Um also eine effektive Formänderung der Zellen zu bewirken, muss eine der beiden Kräfte moduliert werden. Dies kann beispielsweise durch regulatorische Faktoren eingeleitet werden, welche den retrograden Fluss der Aktinfilamente und/oder die Mikrotubuli-basierte Kraft von zytoplasmatischem Dynein direkt oder indirekt beeinflussen (Abb. 4). Es ist noch unklar, welche speziellen Faktoren hierbei eine dominante Rolle spielen.
 die Organisation des Zytoskeletts und Formänderungen in der Entwicklung von Neuronen beeinflussen kann.")
In der momentanen Forschungsarbeit am MPI für molekulare Zellphysiologie studieren diese Wissenschafter der Abteilung Systemische Zellbiologie den Einfluss von Mikrotubuli regulierenden Faktoren auf diesen Prozess. Hierzu verwenden sie einen Forschungsansatz, welcher auf drei Säulen basiert: 1) Hochdurchsatz-Screening, um den relativen Einfluss einzelner Faktoren auf die Neuritenbildung zu bestimmen. 2) Detaillierte Untersuchungen des Zytoskeletts durch Videomikroskopie, um das kollektive Verhalten von Schlüsselkomponenten in der dynamischen Ökologie von lebenden Zellen zu studieren. 3) Mathematische Modellierung, um komplexe und nicht-intuitive Konzepte in der Selbstorganisation der zugrundeliegenden Dynamik des Zytoskeletts zu erfassen. Durch diese Kombination von detaillierter zellbiologischer Forschung und mathematischer Modellierung können die Forscher schrittweise ein besseres Verständnis über die komplexen Vorgänge in sich entwickelnden Neuronen erlangen, welches schließlich neue Wege zur Behandlung von Krankheiten des Nervensystems aufzeigen kann.