Forschungsbericht 2009 - Max-Planck-Institut für Biophysik
Lichtgesteuerte Membrankanäle und Ionenpumpen als Werkzeuge für die Neurowissenschaft
Biophysikalische Chemie (Prof. Dr. Ernst Bamberg)
MPI für Biophysik, Frankfurt am Main
Einführung
Der lichtaktivierbare Ionenkanal Channelrhodopsin2 (ChR2) aus der einzelligen Grünalge Chlamydomonas reinhardtii findet auf Grund seiner einzigartigen Eigenschaft, dass mit ihm elektrisch erregbare Zellen durch Licht depolarisiert und damit angeschaltet werden können, große Beachtung. Obwohl ChR2 weltweit in vielen Labors verwendet wird [1, 2], sind die molekularen Eigenschaften des 2003 entdeckten Proteins wenig erforscht. Daher haben wir uns einerseits verstärkt dem molekularen Mechanismus von ChR2 zugewandt, andererseits wurden von uns auf der Basis der mikrobiellen Rhodopsine neue Werkzeuge zur Lichtsteuerung von Neuronen entwickelt, die einerseits eine erhöhte Lichtempfindlichkeit haben und andererseits eine höhere ortsspezifische Auflösung gestatten. So wurde der Photozyklus des ChR2 analysiert und der Offenzustand des Kanals bestimmten Zwischenprodukten des Photozyklus zugeordnet. Im Einklang mit dem Photozyklus konnte gezeigt werden, dass ChR2 eine Protonenpumpe ist und die Kanaleigenschaften auf ein Leck des Proteins zurückzuführen sind. Die Einzelkanaleigenschaften, wie Leitfähigkeit, Spannungsabhängigkeit und Kinetik, wurden mithilfe einer stationären Rauschanalyse bestimmt.
Molekulare und funktionelle Eigenschaften von Channelrhodopsin2
Wie alle mikrobiellen Rhodopsine besitzt ChR2 als wesentliches Strukturelement sieben membranüberspannende α-Helices. Der Chromophor Retinal ist an ein Lysin der Peptidkette in Helix7 über eine Schiff’sche Base an das Protein gebunden. Um Informationen über das spektrale Verhalten von ChR2 zu erhalten, wurde eine zeitaufgelöste Blitzlichtphotolyse durchgeführt. Für diese Experimente wurde ChR2 heterolog in einem geeigneten Organismus exprimiert; als optimal erwies sich die Hefe Pichia pastoris, aus der nach mehreren Reinigungsschritten das Protein in ausreichenden Mengen erhalten wurde. Die photometrischen Experimente zeigten einen sequenziell zwei Photonen absorbierenden Photozyklus [3]. Der Vergleich mit zeitaufgelösten Strommessungen ergab die Identifizierung eines Offenzustandes von 10 ms Lebensdauer mit einer Rotverschiebung des Absorptionsmaximums von 470 nm im Grundzustand nach 520 nm im Offenzustand. In Zusammenarbeit mit den Arbeitsgruppen von Joachim Heberle, Bielefeld/Berlin, und Georg Nagel, Würzburg, konnte mithilfe der Infrarotspektroskopie gezeigt werden, dass ChR2 beim Übergang vom Grundzustand in den Offenzustand eine vergleichsweise große Konformationsänderung durchläuft, ein Ergebnis, das mit den Ionenkanaleigenschaften im Einklang steht.
Wesentliche Eigenschaften von ChR2, nämlich die Einzelkanalleitfähigkeit und das für die Anwendung wichtige Gleichrichterverhalten des Kanals, wurden mithilfe einer stationären Rauschanalyse untersucht [4]. Die Kenntnis der Einzelkanalleitfähigkeit und der aus der Kinetik des Gesamtstroms bestimmten Lebensdauer der Kanäle erlaubt eine genaue Abschätzung, wie viele Kopien von ChR2 in der zu untersuchenden Wirtszelle aktiv sind - eine Information, die für die Optimierung bei der Expression von ChR2 in Neuronen wichtig ist, wobei es natürlich von Interesse ist, das molekulare Verhalten dieser einzigartigen Ionenkanäle genau zu kennen und zu verstehen. Mit der herkömmlichen patch-clamp-Methode konnten einzelne ChR2-Kanäle nicht bestimmt werden. Daraus wurde geschlossen, dass die Einzelkanalleitfähigkeit extrem niedrig sein muss. In der Tat ergab die Rauschanalyse eine Leitfähigkeit von 40 Femtosiemens. Dieser Wert entspricht einem Einzelkanalstrom von 4 * 10-15 Ampere bei einer Potenzialdifferenz von 100 mV.
Der durch ChR2 induzierte elektrische Strom ist einwärts gerichtet, eine wichtige Voraussetzung für die Anwendung in Nervenzellen, da somit unter physiologischen Bedingungen nur die durch Licht induzierte Depolarisierung auf Grund des Natriumionen Einstroms und die daraus folgende Aktivierung der Zellen möglich ist. Durch Vergleich der Spannungsabhängigkeit der makroskopischen Ströme mit den Einzelkanalströmen von ChR2 ergab sich, dass das Gleichrichterverhalten sowohl auf die Spannungsabhängigkeit der Stromamplitude der Einzelkanäle als auch auf eine veränderte Offenwahrscheinlichkeit zurückzuführen ist. Die Ergebnisse können mit einem einfachen Barrierenmodell für die Ionentranslokation interpretiert werden.
ChR2 weist in den für seine Funktion wichtigen Aminosäure-Sequenzabschnitten eine hohe Homologie mit den entsprechenden Abschnitten der lichtgetriebenen Protonenpumpe Bakteriorhodopsin aus dem Archebakterium Halobacterium salinarum auf. Daher wurde geprüft, ob auch ChR2 als Protonenpumpe agieren kann, nicht zuletzt auch deswegen, weil auf Grund der gewonnenen spektroskopischen Resultate der Photozyklus des ChR2 ebenfalls eine große Ähnlichkeit mit dem des Bakteriorhodopsins aufweist, nämlich die De- und Reprotonierung der Schiff’schen Base, über die der Chromophor Retinal an die Peptidkette (Opsin) gebunden ist. Dieser Prozess ist in der Regel bei mikrobiellen Rhodopsinen mit einem gerichteten Protonentransport verbunden. In der Tat konnte gezeigt werden, dass bei Wegnahme des elektrischen (ΔΨ=0) und des chemischen Gradienten (Δμ=0) ChR2 als auswärts gerichtete Protonenpumpe arbeitet, wobei jedoch die Kanaleigenschaften des Proteins auf eine hohe passive Permeabilität für Kationen zurückzuführen sind, die, wie bereits früher gezeigt, exakt dem elektrochemischen Gradienten folgen.
Neue Werkzeuge für neurobiologische Anwendungen
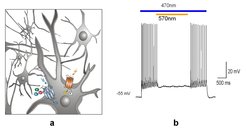
Obwohl der Wildtyp von ChR2 bereits ausgezeichnete Ergebnisse bei vielen neurobiologischen Arbeiten geliefert hat, ist es wünschenswert, verbesserte Versionen mit Hilfe von Punktmutationen oder, in Kombination mit der Cl- Pumpe Halorhodopsin (NphR,), Fusionsproteine, so genannte Tandemmoleküle, zu erzeugen. In einer früheren Arbeit war gezeigt worden, dass NphR durch den Cl- Einwärtsstrom Nervenzellen hyperpolarisiert und damit stilllegt. Tandemmoleküle aus ChR2 und NphR haben den Vorteil, dass eine äquimolare Expression, auch ortsspezifisch, in der Zelle gewährleistet ist, das heißt, Zellen können mithilfe der Tandemmoleküle mit hoher räumlicher Präzision durch ChR2 mit blauem Licht einerseits depolarisiert und damit angeschaltet werden, andererseits mit gelbem Licht durch NphR hyperpolarisiert und damit abgeschaltet werden (Abb. 1).
 und deren Inhibierung durch NphR (gelbes Licht) in kultivierten Hippokampus Zellen. b Aufzeichnung der entsprechenden Aktionspotenziale an der neuronalen Zelle.")
In Experimenten am Gehirn von Nagern ist eine erhöhte Lichtsensitivität dann wünschenswert, wenn nicht nur an der Oberfläche, sondern auch im Innern des Gehirns Untersuchungen vorgenommen werden sollen, da die Lichtdurchlässigkeit des Gehirngewebes äußerst gering ist. Durch Studien an der noch hypothetischen Struktur von ChR2, die bislang nur aus der bekannten Struktur des Bakteriorhodopsins abgeleitet werden konnte, wurde beispielsweise eine ChR2 Mutante spektroskopisch und elektrophysiologisch charakterisiert, bei der sich der Kanal nach einem kurzen Lichtpuls permanent im Offenzustand befindet [5]. Durch geeignete Wahl des Belichtungsprogramms kann somit sogar in bestimmten Zeitintervallen der Kanal ein- bzw. abgeschaltet werden. Durch diese freie Wählbarkeit der Lebensdauer des Kanalzustands ist auch eine Wählbarkeit der Lichtempfindlichkeit gegeben.
Laufende und zukünftige Projekte
Die Abteilung ist in zwei größere Projekte zu neurodegenerativen Krankheiten involviert, bei denen die optogenetischen Werkzeuge angewendet werden. Das erste Projekt beschäftigt sich mit der Anwendung von ChR2 und einiger Mutanten zur Wiederherstellung des Sehens, während im zweiten Projekt nach Alternativen zu der bisher auf Elektroden basierenden Tiefen Hirnstimulation“ bei der Behandlung der Parkinsonschen Krankheit gesucht wird. Ferner bestehen im Bereich der Grundlagenforschung eine Reihe von Kollaborationen im Bereich der Spektroskopie, der Strukturforschung und der Neurobiologie.