Forschungsbericht 2006 - Max-Planck-Institut für biologische Intelligenz (Standort Martinsried)
Zwei Nervenzellen im Direktkontakt - Bewegungssehen durch direkte Verrechnung von optischen Flussfeldern zwischen zwei Hemisphären im Sehzentrum von Fliegen
Neuronale Informationsverarbeitung (Borst) (Prof. Dr. Alexander Borst)
MPI für Neurobiologie, Martinsried
Direkt oder indirekt erzeugte Bildverschiebungen und optische Flussfelder
Bewegungswahrnehmung und –steuerung sind das Ergebnis komplexer Verrechnungen von Signalen im Gehirn, die sowohl von den Augen als auch von der Muskulatur gesendet werden. Augenbewegung, unsere eigene Körperbewegung oder Bewegungen in unserer Umwelt verursachen Bildverschiebungen auf der Netzhaut, die sehr ähnlich oder identisch sind. Das Gehirn kann dennoch diejenigen Bewegungen in der Umgebung von denjenigen unterscheiden, die durch Eigenbewegung erzeugt wurden. Neurobiologen in der Abteilung Neuronale Informationsverarbeitung erforschen die Verrechnung neuronaler Signale am Sehzentrum von Schmeißfliegen. Diese Insekten sind wahre Bewegungskünstler und haben zusätzlich ein überschaubares Sehzentrum mit relativ großen Nervenzellen - geradezu ideale Voraussetzungen, die phantastischen Lösungen der Natur zu studieren und daraus Rückschlüsse auf die Funktion des strukturierten Säugetier- und Menschengehirns zu ziehen oder in technische Anwendungen umzusetzen.
Bewegungen im Raum erzeugen bei Mensch und Tier so genannte optische Flussfelder, die für eine jeweilige Bewegung charakteristisch sind. Bei einer Vorwärtsbewegung beispielsweise fließen die Objekte seitlich vorbei, Objekte frontal vorn wiederum vergrößern sich und weit entfernte Objekte ändern sich in ihrer Abbildung auf der Netzhaut beinahe nicht. Damit die Tiere ihre eigene Bewegung von der bewegten Umwelt unterscheiden und eventuell Kurskorrektur vornehmen können, muss eine Verrechnung der visuellen Information auf höherer Ebene im Sehzentrum des Gehirns stattfinden. Wichtig für die Analyse der Flussfelder ist, dass dabei die Bewegungsinformation von beiden Augen zusammenkommen muss, um das gesamte Flussfeld beurteilen zu können. In einer aktuellen Studie wurde erstmalig die direkte Verschaltung von zwei Nervenzellen nachgewiesen, die aus der jeweils anderen Gehirnhälfte stammten und so die Bewegungssignale von beiden Facettenaugen der Fliege miteinander kombinieren konnten [1].
Neuronale Ausstattung und Erforschung des Schaltplans
Nervenzellen, die optische Flussfelder analysieren können, befinden sich bei der Schmeißfliege in einem höheren Sehzentrum, der so genannten Lobula-Platte. Von diesen so genannten Tangentialzellen existieren pro Gehirnhälfte nur 60, und jede dieser Zellen ist individuell identifizierbar und kommt je Lobula-Platte nur einmal vor. Von diesen 60 Zellen ist eine als H2 bezeichnete Zelle von besonderem Interesse. Diese Zelle zeigt eine starke Präferenz für Rotations-Flussfelder, wie sie bei einer Drehung der Fliege um ihre Hochachse auftreten. Auffallend war, dass diese Zelle zunächst nur auf Bewegungen vor ihrem „eigenen“, dem ipsilateralen Auge, zu reagieren schien, für Bewegungen vor dem anderen Auge, dem kontralateralen, also blind war. Kombinierte man jedoch die ipsilateralen Bewegungsreize mit denen vor dem anderen Auge, war zu erkennen, dass die kontralateralen Bewegungsreize sehr wohl die Reaktionen auf ipsilaterale Bewegungsreize modulierten. Der Präferenz der H2-Zelle für Drehreize liegt also eine nicht-lineare Verrechnung der Bewegungsreize von beiden Augen zu Grunde.
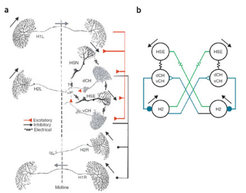
Die Ursachen für diese beobachtete Nicht-Linearität sollten im nächsten Schritt gefunden werden. Dieser bestand darin, die Verschaltung der Tangentialzellen der Lobula-Platte genau zu betrachten. Der sich daraus ergebende Schaltplan basiert auf einer Vielzahl von Experimenten, in denen die Verbindungen zwischen den Zellen innerhalb einer Lobula-Platte und denen zwischen den beiden Hemisphären untersucht wurden. Es ergaben sich letztendlich zwei Wege, auf denen die Bewegungsinformation von der einen Gehirnhälfte die H2-Zelle in der anderen erreichen könnte: Einmal direkt von der so genannten HSE-Zelle, welche mit der H2-Zelle der gegenüberliegenden Hemisphäre elektrisch gekoppelt ist und ein andermal indirekt über die als CH bezeichnete Zelle, die Information über mehrere Stationen von der anderen Gehirnhälfte bekommt und zusätzlich die H2-Zelle, welche auf der gleichen Seite wie sie selbst liegt, chemisch über Synapsen hemmt. Beide Wege sind im Prinzip dazu geeignet, den oben beschriebenen Effekt zu erzielen, also kontralaterale Bewegungsreize auf ipsilaterale Bewegungsreize zu modulieren. Die Frage war nur, welcher von beiden ist der entscheidende?
Um diese Frage zu beantworten, sollten die beiden möglichen Wege selektiv blockiert und anschließend sollte die Rotations-Empfindlichkeit der H2-Zelle getestet werden. Dabei verwendeten die Wissenschaftler die Technik der Laserablation: Füllt man eine einzelne Zelle mit einem fluoreszierenden Farbstoff, der bei starker Beleuchtung für die gefüllte Zelle toxisch wirkt, und bestrahlt das Gehirn anschließend mit Laserlicht der geeigneten Wellenlänge, stirbt diese Zelle innerhalb weniger Minuten. In einer langen Serie dieser technisch sehr schwierigen Experimente gelang der eindeutige Nachweis: Wurde die ipsilaterale CH-Zelle ausgeschaltet, zeigte sich kein Effekt auf die Rotations-Empfindlichkeit der H2-Zelle. Wurde jedoch die kontralaterale HSE-Zelle aus dem Schaltkreis entfernt, war die Rotations-Empfindlichkeit der H2-Zelle verschwunden - nun war sie für Bewegungsreize vor dem anderen Auge tatsächlich blind, egal, ob sie mit ipsilateralen Bewegungsreizen kombiniert war oder nicht. Aus diesen Experimenten konnte ein Schaltplan für 12 der 60 Tangentialzellen im Sehzentrum der Schmeißfliege aufgestellt werden (Abb. 1). Genial an dieser Verschaltung ist ihre Einfachheit: Mit einer einzigen elektrischen Kopplung zweier Zellen aus beiden Gehirnhälften wird eine Zelle selektiv für Rotations-Flussfelder genutzt.
 Darstellung der Verbindungen zwischen den einzelnen beteiligten Neuronen. Rote Dreiecke: aktivierende Verbindungen; schwarze Kreise: hemmende Verbindungen. (b) Vereinfachtes Schema der möglichen Verbindungen zwischen den H2-Zellen beider Hemisphären.")
Die Beantwortung der Frage, ob die Natur bei Säugern auf ähnlich einfache Mechanismen gebaut hat, bleibt abzuwarten. Noch ist die Verschaltung der Nervenzellen in entsprechenden Arealen der Großhirnrinde nicht hinreichend aufgeklärt, um solche Experimente auch dort sinnvoll durchführen zu können. Ob sich vergleichbare experimentelle Effekte zeigen, wenn bei den vielen Milliarden Zellen der Großhirnrinde eine einzelne Zelle herausgenommen wird, ist eher fraglich. Dies bedeutet aber nicht, dass die hier beschriebenen Ergebnisse aus Versuchen mit Schmeißfliegen ohne Konsequenzen für andere Bereiche der Wissenschaft bleiben. So setzen zum Beispiel Ingenieure bei der Entwicklung autonom navigierender Roboter und Fahr-Assistenz-Systemen gern auf einfache und robuste Algorithmen, wie sie die Natur in Insekten längst realisiert hat. Die Mechanismen der optischen Flussfeld-Analyse von Fliegen eignen sich daher hervorragend für eine technische Umsetzung. Daher werden die Max-Planck-Forscher mit Kollegen von der Technischen Universität München im Rahmen zweier vom Bundesministerium für Bildung und Forschung (BMBF) geförderter Projekte (Bernstein Zentrum München und Cognition in Technical Systems - CoTeSys) die nächsten Jahre verstärkt daran arbeiten. Darüber hinaus ist die Abteilung Neuronale Informationsverarbeitung an der kürzlich in die Exzellenzinitiative aufgenommenen Graduiertenschule School of Systemic Neurosciences der Münchner Ludwig-Maximilians-Universität beteiligt.