Forschungsbericht 2020 - Max-Planck-Institut für biologische Intelligenz (Standort Martinsried)
Wie rechnen Nervenzellen?
Warum die Erforschung des Bewegungsehens in der Fliege?
 gliedert sich in ein Thorakal- und ein Kopfganglion. Im Kopfganglion (links) ist der optische Lobus (rot) besonders ausgeprägt. B Aufbau des optischen Lobus. C Auf Bewegung eines Streifens reagieren die Lichtsinneszellen unabhängig von der Richtung (oben). Zellen der Lobulaplatte hingegen zeigen richtungs-abhängige Signale (unten).")
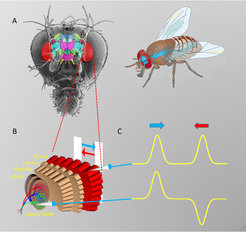
Informationen darüber zu erhalten, in welche Richtung sich ein Objekt bewegt, kann für das Überleben eines Organismus entscheidend sein – das gilt auch für unser Untersuchungsobjekt, die Fliege Drosophila. Um die allgemeine Frage zu beantworten, wie Nervenzellen rechnen, ist ihr Bewegungssehen in besonderem Maße geeignet. Dem Bewegungssehen liegt eine einfache Art der Berechnung zugrunde, welche theoretisch sehr gut verstanden ist. Dazu kommt, dass das Fliegengehirn im Vergleich zum menschlichen Gehirn mit seinen circa Einhundert Milliarden Nervenzellen lediglich aus etwa 100.000 Nervenzellen besteht (Abb.1, A). Dies erleichtert seine Analyse entscheidend. Bei der Fruchtfliege Drosophila stehen zudem genetische Methoden zur Verfügung, Nervenzellen gezielt, also zelltyp-spezifisch, zu aktivieren und zu inaktivieren. Weiterhin ist es möglich, Proteine in die Zellen einzubringen, die ihre Fluoreszenz erhöhen, wenn die Nervenzellen aktiv sind. Dadurch lässt sich deren Aktivität im Mikroskop messen.
Das Sehsystem der Fliege
Die Lichtsinneszellen der Fliege befinden sich in den einzelnen Facetten ihrer großen Augen. Der dahinterliegende ‚optische Lobus‘ besteht aus vier Schichten von Nervenzellen, welche sich in einzelne Säulen (Kolumnen) gliedern (Abb.1, B). Diese Kolumnen entsprechen in ihrer Anzahl und Anordnung genau den Facetten des Auges (Drosophila besitzt ungefähr 750 Facetten pro Auge). Bewegt sich ein Objekt nach rechts und wieder zurück, antwortet eine einzelne Lichtsinneszelle beide Male gleich (Abb.1, C, oben): die Information über die Bewegungsrichtung ist also nicht aus den Signalen einer einzelnen Lichtsinneszelle ablesbar. Anders die großen Tangentialzellen der Lobulaplatte: Sie reagieren auf Bewegung in eine Richtung mit einer elektrischen Erregung, auf Bewegung in die andere Richtung mit einer Hemmung (Abb.1, C, unten). Innerhalb weniger Verknüpfungsstellen wird also aus einem richtungs-unabhängigen Signal ein richtungs-abhängiges Signal errechnet. Doch wo genau geschieht das?
Fluoreszierende Proteine bringen Licht ins Dunkel der Signalverarbeitung
. C. Mikroskop-Aufnahme eines Horizontalschnitts durch den optischen Lobus. Die T4 und T5 Zellen sind mit einem Aktivitäts-abhängigen Fluoreszenz-Farbstoff grün markiert. Gelb umrandetet ist der Ausschnitt, der in D gezeigt ist. D. Je nach ihrer Schicht in der Lobula Platte reagieren die T4 und T5 Zellen auf Bewegung entlang einer der vier Hauptrichtungen. E. Elektrisches Ersatzschaltbild der T4 Zellmembran zusammen mit den Antworteigenschaften (oben) von drei Eingangszellen. Wichtig dabei ist, dass Mi9 durch einen hellen Punkt gehemmt (blau), Mi1 und Mi4 aber erregt werden (rot). Die Eingangszellen regulieren mit ihrer Aktivität unterschiedliche Leitfähigkeiten in der Zellmembran von T4. F. Antworten des T4-Modells (links) im Vergleich zu echten T4 Zellen (rechts) auf verschiedene Stimuli. Oben: Ein periodisches Streifenmuster bewegt sich mit verschiedenen Geschwindigkeiten (ausgedrückt durch die Kontrast-Frequenz, d.h. Geschwindigkeit geteilt durch Muster-Wellenlänge) von links nach rechts (‚preferred direction‘, PD, in blau) und von rechts nach links (‚null direction‘, ND, in rot). Unten: Das Streifenmuster bewegt sich mit 30 Grad/s in 12 verschiedene Richtungen. Die Antworten sind in Polarkoordinaten dargestellt.")
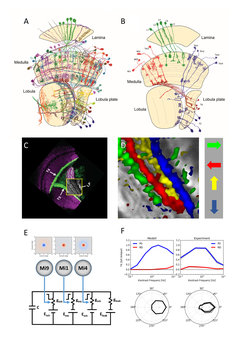
Jede Kolumne des optischen Lobus enthält etwa 100 verschiedene Nervenzell-Typen (Abb.2 A, B). Diese sind schon vor vielen Jahrzehnten anatomisch gut beschrieben worden, ihre Signale konnten jedoch, bedingt durch die winzige Größe der Zellen, nicht mit Mikro-Elektroden gemessen werden. Somit blieben die Antworteigenschaften der verschiedenen Zelltypen lange im Dunkeln. Ihre genaue Beschreibung gelang erst durch die Verwendung aktivitäts-abhängiger Fluoreszenzproteine. Wie sich herausstellte, sind die es die sogenannten T4- und T5-Zellen, die als erste Zellen in der Verarbeitungsbahn richtungsabhängig auf Bewegung antworten. In jeder Kolumne existieren vier T4- und vier T5-Zellen. Während T4-Zellen bevorzugt auf die Bewegung von hellen Kanten reagieren, sprechen T5-Zellen vorzugsweise auf die Bewegung von dunklen Kanten an. Von beiden Zelltypen gibt es vier funktionelle Gruppen (Abb. 2; C,D): Eine Gruppe von T4/T5-Zellen reagiert auf Bewegung von vorne nach hinten (grün dargestellt), eine andere auf Bewegung von hinten nach vorne (rot), eine dritte Gruppe auf Aufwärtsbewegung (gelb), eine vierte Gruppe auf Abwärtsbewegung (blau). Je nach ihrer Vorzugsrichtung ragen die Fortsätze der T4/T5-Zellen in eine der vier Lagen der Lobulaplatte, wo sie die großen Tangentialzellen kontaktieren. Wie errechnen aber die verschiedenen Untergruppen von T4/T5-Zellen die jeweilige Bewegungsrichtung? Dies ist vor allem für T4-Zellen schon recht gut verstanden.
Zelluläre Mechanismen der Richtungs-Selektivität
T4-Zellen erhalten ihre Eingangssignale unter anderem von den sogenannten Mi1-, Mi4- und Mi9-Zellen. Interessanterweise folgt die räumliche Anordnung der Zellkontakte einem Prinzip: T4-Zellen, die für Bewegung von vorne nach hinten empfindlich sind, erhalten das Signal von Mi9 auf der linken Seite, von Mi1 in der Mitte und von Mi4 auf der rechten Seite des Dendriten (Abb.2, E). T4-Zellen, die auf Bewegung von hinten nach vorne reagieren, erhalten ihren Input in umgekehrter Reihenfolge.
Aus verschiedenen Studien sind sowohl die Neurotransmitter als auch deren Rezeptoren in den T4 Zellen bekannt. Mit einem elektrischen Ersatzschaltbild der T4-Zellmembran lässt sich nun aus den Signalen der Eingangszellen das Signal der T4-Zelle berechnen. Wie entsprechende Computersimulationen zeigen, stimmen die Antworten des T4-Modells recht gut mit den experimentell an T4-Zellen gewonnenen Daten überein (Abb.2, F). Die Zukunft wird zeigen, ob unser Modell der experimentellen Überprüfung umfassend standhält.
Zusammenfassung und Ausblick
Das Sehsystem der Fliege hat durch seinen regelmäßigen, nahezu kristallinen Aufbau seit jeher das Interesse der Forschung geweckt. In den vergangenen Jahren sind zu dem Wissen über die Anzahl und Anatomie aller Nervenzelltypen auch die Kenntnis ihrer genauen Verknüpfung, das sogenannte ‚Connectome‘, hinzugekommen. Außerdem kennt man mittlerweile von fast allen Zelltypen sowohl deren Neurotransmitter als auch viele weitere, zellspezifische Proteine. Dies bringt die Forschung in die einmalige Lage, allgemeine Fragen zur Funktion des Nervensystems zu beantworten, die sich an kaum einem anderen Modellsystem beantworten lassen. Eine dieser Fragen ist die, wie Nervenzellen die Bewegungsrichtung errechnen. Die Antwort auf diese Frage scheint zum Greifen nahe und wird in naher Zukunft dazu führen, dass mithilfe eines einfachen Modellsystems im Detail verstehen, wie das Nervensystem auf der Ebene einzelner Nervenzellen und deren präziser Verknüpfung neuronale Berechnungen durchführt - Berechnungen, die für das Überleben von Individuen von entscheidender Bedeutung sind.
Literaturhinweise
Nature 500, 212-216 (2013)
PLoS Comput Biol 14: e1006240 (2018)