Forschungsbericht 2010 - Max-Planck-Institut für Infektionsbiologie
Tuberkulose: Forschung über eine vergessene, aber nicht vergangene Krankheit
Max-Planck-Institut für Infektionsbiologie, Berlin
Einleitung
Zwar stehen uns Medikamente zur Heilung der Tuberkulose (TB), Diagnostika zur Erkennung der Erkrankung und ein Impfstoff zur Verhinderung der schlimmsten Formen der TB des Kleinkinds zur Verfügung [1]. Dies reicht aber nicht aus:
- Die Diagnose der TB wird in denjenigen Ländern, die von der Krankheit am meisten betroffen sind, lediglich in der Hälfte der Fälle richtig gestellt.
- Die Behandlung der TB bedarf der Gabe von drei bis vier verschiedenen Medikamenten über sechs Monate und länger. Dies führt häufig zum vorzeitigen Abbruch der Medikamenten-Einnahme. Heute sind bereits 50 Millionen Menschen mit multiresistenten (multi-drug resistant, MDR) Mycobacterium tuberculosis (Mtb) Erregern infiziert, von denen jährlich eine halbe Million Menschen erkranken. Die Multiresistenzen nehmen besonders in Osteuropa in erschreckendem Maße zu, wo sie inzwischen in vielen Regionen deutlich mehr als 10% ausmachen. Noch bedrohlicher sind die so genannten extensiv resistenten TB-Fälle (XDR-TB), die bereits aus 58 Ländern gemeldet wurden. Diese Erreger sind gegen alle herkömmlichen Medikamente resistent.
- Der verfügbare Impfstoff bacille Calmette Guérin (BCG) schützt zwar gegen heftig verlaufende Krankheitsverläufe bei Kleinkindern; gegen die am weitesten verbreitete Form der Erkrankung, die Lungentuberkulose bei Erwachsenen, ist BCG jedoch nicht wirksam.
. Die Abbildung zeigt in rot die Anzahl der Infektionen, der Krankheitsausbrüche und der Todesfälle, die durch Mycobacterium tuberculosis (Mtb) hervorgerufen werden. Weiterhin zeigt die Abbildung die Bedrohung durch Koinfektion mit dem humanen Immundefizienz-Virus (HIV) und durch multiresistente TB (MDR-TB).")
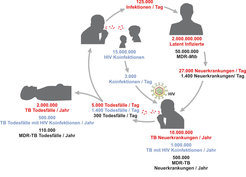
Somit ist die TB entgegen der häufig geäußerten Meinung, dass diese Erkrankung der Vergangenheit angehöre, auch heute noch eine ernst zu nehmende Bedrohung. Die unter anderem von der Weltgesundheitsorganisation (WHO) getragene Organisation Stop TB Partnership hat sich zum Ziel gesetzt, die TB bis zum Jahr 2050 zu eliminieren, das heißt auf weniger als einen Fall pro einer Million Einwohner zu reduzieren [1, 2]. Dieses ehrgeizige Ziel kann nur erreicht werden, wenn uns neue Impfstoffe, neue Medikamente und bessere Diagnostika zur Verfügung stehen. Unsere Arbeitsgruppe beschäftigt sich unter anderem mit der Entwicklung eines wirksamen und sicheren Impfstoffs gegen TB und mit der Charakterisierung von Biomarkern zur Unterscheidung von latent infizierten Gesunden und aktiv Erkrankten sowie zur Abschätzung des Risikos einer aktiven TB in Infizierten. Von den geschätzten 2 Milliarden Menschen, die mit Mtb infiziert sind, hat nämlich nur jeder Zehnte zu befürchten, dass er eines Tages an TB erkranken wird. Allerdings sind dies derzeit immer noch 10 Millionen neue Fälle jährlich, von denen jeder Fünfte, also 2 Millionen, stirbt. Abbildung 1 fasst die wichtigsten Morbiditäts- und Mortalitätsdaten zusammen.
Unsere Forschung geht von der Annahme aus, dass eine Aufschlüsselung des komplexen Wechselspiels zwischen dem Erreger Mtb und seinen Wirtszellen, insbesondere den Zellen des Immunsystems, die Grundlagen für bessere Interventionsmaßnahmen legen kann [3]. Dieser heute häufig als Translationsforschung bezeichnete Ansatz hat bereits zu einem Impfstoff, der die erste klinische Überprüfung bestanden hat, verholfen und den Beweis für das Prinzip erbracht, dass latente Infektion und aktive TB mithilfe von Biomarkern unterschieden werden können.
Mechanismen der Infektabwehr
Eines der bemerkenswerten Charakteristika der TB ist der große Anteil an latent infizierten Personen, die nie an TB erkranken. Bei diesen Personen ist die körpereigene Immunantwort in der Lage, die Infektion einzudämmen; allerdings gelingt es ihr nicht, den Erreger zu eliminieren. Dies führt zu einer latenten Infektion. Mtb besitzt die Fähigkeit, in körpereigenen Abwehrzellen zu überleben. Dies sind die mononukleären Phagozyten, insbesondere die Gewebsmakrophagen. Während die meisten Mikroben von diesen professionellen Fresszellen abgetötet werden, kann Mtb überleben. Nach ihrer Aufnahme in die Fresszellen gelangen die Keime in das Phagosom, das heranreift und mit dem Lysosom verschmilzt. Im Phagolysosom werden die Erreger gewöhnlich eliminiert. Dieser Prozess wird von Mtb unterbunden (siehe auch Abb. 3a). Das Phagosom verharrt in einem frühen Stadium, in dem Mtb persistieren kann. Aufgrund seines intrazellulären Lebens ist der Erreger weitgehend vor Angriffen durch Antikörper geschützt. Bestandteile des Erregers werden aber im Makrophagen prozessiert und von Referenzstrukturen präsentiert. Dies führt zur Aktivierung von T-Lymphozyten, die die infizierten Makrophagen spezifisch erkennen. Nach ihrer Aktivierung produzieren die T-Lymphozyten unterschiedliche Zytokine, die wiederum die Makrophagen aktivieren, sodass diese den Erreger in Schach halten, allerdings nicht eliminieren. Wichtige Botenstoffe sind Interferon-γ (IFN-γ) und Tumor-Nekrose-Faktor (TNF).
In den meisten Fällen findet die Auseinandersetzung in der Lunge statt, denn der Erreger wird fast ausschließlich über Aerosole übertragen. Um dem Angriff der Immunzellen zu widerstehen, verfällt der Erreger in eine Art „Winterschlaf“, in dem er seinen Stoffwechsel auf ein Minimum reduziert und somit gegenüber der Immunabwehr resistent wird. Dieses aktive Wechselspiel wird erst dann gestört, wenn das Immunsystem geschwächt wird. Dann reichen die körpereigenen Abwehrkräfte zur Erregerkontrolle nicht mehr aus, Mtb wird aufgeweckt und vermehrt sich und eine aktive TB bricht aus. Über die Blutkapillaren gelangen die Erreger in den Brutkreislauf, werden über den Körper verbreitet und befallen andere Organe. Auch den Alveolarraum erreichen jetzt die Erreger. Der Patient ist ansteckend, da er beim Husten die Keime in die Umgebung versprüht. Die kleinen Tröpfchen enthalten einige Mtb-Organismen, die von Menschen in der Umgebung eingeatmet werden.
Da sich Mtb bevorzugt im Phagosom des Makrophagen aufhält, können Erreger-Bestandteile in den so genannten MHC-Klasse II Präsentationsweg eingeschleust werden. Bei diesem Weg übernehmen Genprodukte des Haupthistokompatibilitätskomplexes (MHC für major histocompatibility complex) der Klasse II die Präsentation von Peptiden. Dies führt zur bevorzugten Stimulierung von CD4 T-Lymphozyten, welche die antigenen Peptide im Kontext von MHC II erkennen. Diese CD4 T-Zellen üben bevorzugt Helferfunktionen aus.
Drei unterschiedliche Klassen von T-Helferzellen (Th) sind bekannt:
- Th1-Zellen, die über IFN-γ, TNF und andere Faktoren Makrophagen und über Interleukin (IL)-2 T-Zellen stimulieren;
- Th2-Zellen, die über IL-4 und IL-5 B-Zellen zur Antikörperproduktion anregen und an der Abwehr von Wurminfektionen beteiligt sind;
- Th17-Zellen, die über die Produktion von IL-17 Neutrophile (dies sind kurzlebige professionelle Phagozyten) aktivieren.
Entscheidend für die Infektabwehr bei der TB sind die Th1-Zellen. Neuere Befunde weisen auf die zusätzliche Beteiligung von Th17-Zellen hin. Th2-Zellen scheinen dagegen keine oder eine eher schädliche Rolle auszuüben. Lange glaubte man, dass Mtb sich nur im Phagosom aufhalte. Neuere Arbeiten haben jedoch Hinweise dafür erbracht, dass der Erreger auch in das Zytoplasma entweichen kann (s. auch Abb. 3b) [4]. Dort werden Erregerbestandteile in den MHC Klasse-I-Weg eingeschleust. Dies führt zur Stimulierung von CD8 T-Lymphozyten, welche kurze Peptide im Kontext von MHC I erkennen. Die CD8 T-Zellen üben in erster Linie zytolytische Funktionen aus. Über die Produktion von Perforinen, Granulysinen und Gramzymen werden nicht nur körpereigene Zielzellen abgetötet, sondern auch Mtb-Zellen zerstört. Eine Beteiligung von CD8 T-Zellen an der Abwehr der TB war schon vorher bekannt. Die Erreger-Egression in das Zytoplasma erlaubt nun eine schlüssige Erklärung des Vorgangs. Untersuchungen aus unserem Labor haben einen zweiten Weg der CD8 T-Zellstimulierung bei der TB aufgeklärt [5]. Beide Wege schließen sich nicht aus und finden mit großer Wahrscheinlichkeit parallel statt. Der von uns aufgeklärte Weg beginnt mit der Apoptose der Mtb befallenen Makrophagen (s. auch Abb. 4). Dieser induzierte Zelltod führt zur Bildung von apoptotischen Vesikeln, in denen sich Bestandteile von Mtb angesammelt haben. Diese apoptotischen Vesikel werden von dendritischen Zellen (dies sind professionelle antigenpräsentierende Zellen) aufgenommen, die dann CD4 und CD8 T-Zellen antigenspezifisch stimulieren.
Das Granulom – Gewebskorrelat von Schutz und Schaden
Das Granulom bildet das histologische Korrelat des Schutzes wie auch des Schadens bei der TB. Solide Granulome sind in der Lage, Mtb einzudämmen, während das nekrotische und mehr noch das verkäste Granulom in erster Linie pathologische Schäden bewirkt. Das Granulom bei der TB des Menschen ist das Resultat einer organisierten Gewebsreaktion. Bei latent Infizierten wird der Erreger in einem soliden Granulom eingedämmt. Dieses besteht hauptsächlich aus Makrophagen und T-Lymphozyten in strukturierter Konstellation. Dies erleichtert die Koordination zwischen diesen beiden Zelltypen, die für eine effektive Erregerkontrolle benötigt wird. Bei Krankheitsausbruch entwickeln sich zuerst nekrotische Granulome, deren Zentren aus nekrotisierenden und zerfallenden Wirtszellen bestehen. Die Erreger können sich nun in dieser festen Masse vermehren. Im Verlauf der Krankheit verkäst der Bereich immer mehr und die Erreger vermehren sich unkontrolliert. Die geordnete Struktur des Granuloms geht verloren. Das Zentrum des soliden und auch des nekrotischen Granuloms ist hypoxisch, das heißt sauerstoffarm, und dieser Zustand trägt zur Kontrolle des Erregers bei, der Sauerstoff zum Überleben benötigt. Um in diesem hypoxischen Milieu zu persistieren, schaltet Mtb seinen Stoffwechsel weitgehend ab: Der Erreger befindet sich in einem dormanten Zustand.

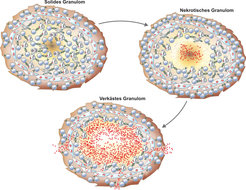
Die Läsionen der experimentellen TB der Maus sind kaum strukturiert und weisen insbesondere kein hypoxisches Zentrum auf. Es gelang, ein Mäusemodell der TB zu entwickeln, bei dem solide, nektrotisierende und verkäsende Granulome nacheinander entstehen [6]. Hierzu wurden Mäuse verwendet, deren Abwehrzellen keine Stickstoffmetabolite bilden können. Bei der Maus übernehmen diese hochreaktiven Metabolite entscheidende Abwehrfunktionen. Beim Menschen werden sie nur in geringem Maße gebildet. Weiterhin ermöglichte der gewählte intradermale Infektionsweg eine rasche Aktivierung von T-Lymphozyten. So gelang es, im Mäusesystem die Ausbildung von Granulomen zu stimulieren, die einerseits ähnlich den humanen Granulomen eine geordnete Struktur aufweisen, andererseits aufgrund der fehlenden reaktiven Stickstoffmetabolite nur ein eingeschränktes Abwehrarsenal mobilisieren können. In diesen Mäusen entwickeln sich Granulome, die den soliden Granulomen bei der Human-TB sehr ähnlich sind. Neutralisierung des für die Makrophagen-Aktivierung zentralen Zytokins IFN-γ führte zur Nekrotisierung und schließlich zur Verkäsung der Granulome. Somit konnte das gesamte Spektrum der Granulom-Entwicklung bei der Human-TB vom protektiven zum pathologischen Reaktionsherd in der Maus nachgebildet werden. Dieses Mausmodell erlaubte auch die Identifizierung eines wesentlichen Effektormoleküls der Abwehr: Cathepsin-G. Dieses aggressive Effektormolekül wird wiederum von Serpin-b3 kontrolliert. Abhängig vom fein abgestimmten Gleichgewicht wirkt Cathepsin-G entweder schützend oder schädlich. Inwieweit dieses molekulare Wechselspiel auch an der Entscheidung über Schutz oder Schaden bei der Human-TB beteiligt ist, wird derzeit untersucht. Abbildung 2 zeigt die verschiedenen Stadien des Granuloms bei der Human-TB und dem neu entwickelten Mausmodell.
Das immunologische Frühwarnsystem
Die angeborene Immunität spielt bei der Erkennung und Kontrolle des Erregers eine wesentliche Rolle. Zahlreiche Bestandteile von Mtb werden von mustererkennenden Rezeptoren unterschiedlichen Typs als spezifische Muster wahrgenommen. Toll-ähnliche Rezeptoren (Toll-like receptors, TLR) erkennen bestimmte Glykolipide und Oligonukleotide; Nod-ähnliche Rezeptoren (Nod-like receptors, NLR) reagieren unter anderem auf Muramyldipeptid; Lektin-Rezeptoren vom Typ C (C-type lectin receptors) sind spezifisch für bestimmte Zucker und Wachse. Dieses breite Erkennungsspektrum ermöglicht eine hohe Kompensationsfähigkeit des Systems, das heißt das Fehlen einzelner molekularer Muster bleibt ohne sichtbare Konsequenz. Die Signale, die von den mustererkennenden Rezeptoren ausgehen, werden von Adaptoren gesammelt und so verarbeitet, dass über neu geordnete Signale geeignete Effektorfunktionen gezielt stimuliert werden. Card 9 ist ein solches Adaptermolekül. Neuere Arbeiten aus unserer Gruppe haben die zentrale Rolle von Card 9 bei der TB aufgezeigt [7]. Mäuse mit einem Defekt in Card 9 sind äußerst empfänglich gegenüber einer Infektion mit Mtb. In diesen Tieren kommt es zu einem starken Einstrom von Neutrophilen in den Infektionsherd. Dies löst eine markante Schädigung der Lunge aus. Grund für die unkontrollierte Entzündungsreaktion ist die weitgehend eingeschränkte Produktion des Botenstoffs IL-10 durch Neutrophile in Card 9 defizienten Tieren. Diese Untersuchungen legen erstens eine schädliche Rolle von Neutrophilen bei der TB nahe und zeigen zweitens einen direkten Rückkopplungsmechanismus auf, bei dem die schädigende Zelle ihren eigenen Inhibitor bildet.
Translationsforschung I: Entwicklung eines Impfstoffs gegen TB
 verbleiben im Phagosom, dessen Reifung sie verhindern. Das frühe Phagosom bildet einen geeigneten Lebensraum für beide Mikroben. Es kommt primär zur Stimulierung von CD4 T-Zellen. B. Mtb kann in das Zytoplasma entweichen; es kommt zur Stimulierung von CD8 T-Zellen. Dagegen verbleibt BCG im Phagosom und kann lediglich CD4 T-Zellen stimulieren. C. Listeria monocytogenes sezerniert Listeriolysin (Hly), das bei einem pH 5,5 Membranporen bilden kann. Auf diese Weise entweicht L. monocytogenes in das Zytoplasma. Es kommt zur Stimulierung von CD4 und CD8 T-Zellen. D. Durch chromosomale Integration des Hly-Gens und Deletion des Urease-Gens (ΔureC) wurde der Impfstamm r-BCG ΔureC:Hly mit porenbildender Aktivität konstruiert. Antigene gelangen in das Zytosol der Zelle und CD4 und CD8 T-Zellen werden stimuliert.")
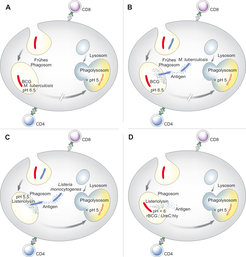
Derzeit suchen mehrere Laboratorien intensiv nach einem neuen TB-Impfstoff. Unsere Strategie zur Entwicklung eines TB-Impfstoffs basiert auf unseren Einblicken in die schützende Immunantwort gegen TB. Die folgenden Befunde waren von besonderer Bedeutung:
- Neben CD4 T-Zellen sind auch CD8 T-Zellen am Schutz gegen TB beteiligt.
- Mtb ist in der Lage, CD8 T-Zellen neben CD4 T-Zellen zu stimulieren.
- Der vorhandene Impfstoff BCG stimuliert lediglich CD4 T-Zellen, aber nicht CD8 T-Zellen.
Das intrazelluläre Bakterium Listeria monocytogenes perforiert die Membran des Phagosoms mithilfe von Listeriolysin (Hly). Dies ermöglicht die Egression des Erregers in das Zytoplasma, wo seine Antigene in den MHC I-Weg eingeschleust werden. Abbildung 3 gibt eine vereinfachte Darstellung der Mechanismen, die dem Entwicklungskonzept zu einem neuen TB-Impfstoffs zugrunde liegen.
 aufgenommen. Durch Perforation der phagosomalen Membran gelangen Antigene in das Zytoplasma. Die Beladung von MHC II-Molekülen im Phagosom und von MHC I-Molekülen im Zytoplasma führt zur Stimulierung von CD4 und CD8 T-Zellen. Makrophagen (Mφ) nehmen r-BCG ΔureC:Hly-Keime auf. Die Perforation der phagosomalen Membran führt zur Apoptose der Wirtszelle. Apoptotische Vesikel, die Antigene des Impfstamms enthalten, werden gebildet und anschließend von DC aufgenommen. Über diesen Prozess, der als \"Crosspriming\" bezeichnet wird, werden mit hoher Effizienz antigenspezifische CD4 und CD8 T-Zellen stimuliert.")
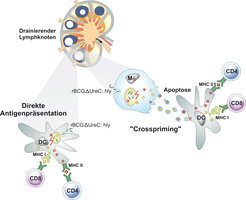
Das Hly kodierende Gen wurde in den Chromosomenring von BCG eingepflanzt. Da Hly zu seiner biologischen Aktivität einen saueren pH 5,5 benötigt, wurde weiterhin ein Gen, das eine Urease (Ure C) kodiert, deletiert. Das Konstrukt erhielt die Bezeichnung r-BCG ΔureC:Hly [8]. Die Fähigkeit des r-BCG ΔureC:Hly Impfstoffs, das Phagosom zu neutralisieren, ist auf diese Weise eingeschränkt. Es kommt zum pH-Abfall und Hly kann seine biologische Aktivität entfalten. Das r-BCG ΔureC:Hly Konstrukt selbst entweicht zwar nicht in das Zytoplasma der Wirtszelle, seine Bestandteile können aber dorthin gelangen und so antigene Peptide auf MHC I-Moleküle geladen werden. Es kommt zur Stimulierung von CD8 T-Zellen in Ergänzung zu CD4 T-Zellen. Weitere Untersuchungen unserer Gruppe zeigten, dass der r-BCG ΔureC:Hly Impfstamm verstärkt Apoptose in den besiedelten Makrophagen hervorruft. Dieser induzierte Zelltod führt zur Bildung von apoptotischen Vesikeln, die Bestandteile des Impfstoffs enthalten. Die Vesikel werden von dendritischen Zellen aufgenommen und die enthaltenen Peptide sowohl CD4 als auch CD8 T-Zellen angeboten. Dieser als crosspriming bezeichnete Präsentationsweg ist äußerst effizient. Neuere Untersuchungen haben Hinweise dafür erbracht, dass über crosspriming auch TH-17 Zellen stimuliert werden. Somit ist die durch r-BCG ΔureC:Hly hervorgerufene Apoptose ein zentraler Schritt, der den stärkeren Impfschutz gegen TB zwanglos erklären kann. Abbildung 4 beschreibt summarisch die Mechanismen, die der verbesserten Impfstoffaktivität des r-BCG ΔureC:Hly Konstrukts zugrunde liegen. In der Tat bewirkt r-BCG ΔureC:Hly einen bis zu hundertfach höheren Schutz gegen Mtb. Der neue Impfstoff erwies sich im Tiermodell außerdem als sicher und wurde insbesondere von immundefizienten Mäusen sehr gut toleriert. Der Impfstoff wurde an die Vakzine Projekt Management (VPM) lizenziert, die die Zulassung für die klinische Überprüfung erhielt, die der Impfstoff uneingeschränkt bestand. Das heißt, er ist auch im Menschen sicher und immunogen. Derzeit unterzieht sich der Impfstoff der klinischen Überprüfung in einem TB-endemischen Gebiet Südafrikas.
Translationsforschung II: Die Entwicklung einer Biosignatur
Die klinische Prüfung eines TB-Impfstoffs muss zahlreiche aufwendige Schritte durchlaufen. Nach der Überprüfung auf Sicherheit und Wirksamkeit erfolgt die Überprüfung der Schutzwirkung in einem abgestuften Prozess. Bis ein Impfstoff die letzte Phase III der klinischen Testung bestanden hat, wurden mehrere zehntausend Probanden geimpft und zahlreiche Jahre sind vergangen. Eine Beschleunigung dieses aufwendigen Prozesses wäre äußerst wünschenswert [1]. Hierzu können Biomarker beitragen, die während einer frühen Phase der klinischen Überprüfung Informationen über die Effektivität eines Impfstoffkandidaten liefern. Wahrscheinlich werden mehrere unterschiedliche Biomarker benötigt, die in einer Biosignatur zusammengefasst werden. Insbesondere eine Biosignatur, die den klinischen Endpunkt (das heißt TB-Erkrankung beziehungsweise Schutz dagegen) voraussagen kann, wäre äußerst hilfreich. Biosignaturen sind für zahlreiche Zwecke nützlich:
- Eine Biosignatur erlaubt die Differential-Diagnose von latent infizierten Gesunden und Patienten mit aktiver TB. Dies kann zu einem diagnostischen Test führen, der unter einfachsten Bedingungen eine eindeutige Diagnose liefert (sogenannter Point of Care Test).
- Eine Biosignatur kann entscheidende Beiträge zum Verständnis der Wechselwirkungen zwischen dem Erreger Mtb und der körpereigenen Abwehr leisten. Im Sinne einer reversen Translation können Biomarker identifiziert werden, deren Rolle dann im Labor durch hypothesengetriebene Forschung aufgeklärt wird.
- Wie bereits erwähnt, können Biosignaturen zum Monitoring klinischer Studien mit Impfstoffkandidaten, aber auch mit neuen Medikamenten herangezogen werden.
- Das ehrgeizigste Ziel wäre eine Biosignatur, die unter den latent infizierten Gesunden diejenigen identifiziert, die zu einem späteren Zeitpunkt an aktiver TB erkranken werden. Diese Biosignaturen erlauben somit eine Prognose des Krankheitsrisikos und erfüllen damit die Kriterien von Korrelaten des Schutzes.
Unsere Gruppe hat in Zusammenarbeit mit zahlreichen Partnern auf dem afrikanischen Kontinent mit der Charakterisierung von Biosignaturen begonnen. Die ersten Arbeiten beruhen auf dem Vergleich von i.) Patienten mit aktiver TB, ii.) latent Mtb infizierten gesunden Probanden und iii.) mit Mtb nicht infizierten gesunden Kontrollen. Es zeigte sich, dass eine handvoll Biomarker zur Unterscheidung dieser drei Gruppen ausreicht [9]. Entscheidende Marker sind:
- Fcγ-Rezeptor, der Antikörper über deren Fc-Stück bindet.
- Laktoferin, welches beim Wettbewerb zwischen Mtb und Wirtszellen um Eisen eine entscheidende Rolle spielt.
- Guanylat-bindendes Protein 5, ein Mitglied großer GTP-bindender Proteine, das an der Signaltransduktion nach Stimulierung mit Zytokinen wie IFN-γ und TNF beteiligt ist.
- Granzym-A, ein zytolytisches Molekül, das an der Zielzell-Lyse durch CD8 T-Zellen mitwirkt.
Diese vier Biomarker erlaubten in zwei Studien mit hoher Spezifität und Sensitivität die Unterscheidung zwischen Patienten mit aktiver TB und latent Mtb-infizierten beziehungsweise nicht infizierten Gesunden. Sensitivität und Spezifität wurden weiter erhöht, wenn zusätzlich die antimikriobiellen Moleküle Defensin-α1 und α4 in die Signatur eingebracht wurden. Die Unterscheidung zwischen aktiver TB und latenter Infektion war also mithilfe dieser handgeschneiderten Biosignatur gelungen.
Für ein besseres Verständnis der Immunantwort, die den Erreger während der latenten Infektion in Schach hält, wurden die Genexpressionsprofile selektionierter T-Zellen erstellt. Wir identifizierten Signaltransduktionsmoleküle, die an der T-Zellregulation entscheidend beteiligt sind [10]. Hierzu gehörten insbesondere Mitglieder des JAK/STAT-Signaltransduktionswegs (JAK = Janus Kinase; STAT = signal transducers and activators of transcription). Eine genauere Analyse zeigte, dass die differenziell regulierten Genprodukte in engem Wechselspiel stehen. Wir vermuten daher, dass Störungen dieser Transduktionswege am Übergang von latenter Infektion zu aktiver TB mitwirken. Genexpressionsprofile von Blutzellen der Probanden der ersten klinischen Studie mit unserem Impfstoffkandidaten r-BCG ΔureC:Hly wurden ebenfalls ermittelt. Zahlreiche differenziell exprimierte Gene konnten identifiziert werden. Die meisten waren nach Impfung mit BCG als auch mit r-BCG ΔureC:Hly gleichermaßen aktiviert. Einige Gene der angeborenen Immunität zeigten ein differenzielles Expresssionsmuster zwischen BCG- und r-BCG ΔureC:Hly.
Weitere Biomarkeruntersuchungen konzentrieren sich auf Stoffwechselprodukte. Diese Metabolomanalysen erwiesen sich ebenfalls als äußerst aufschlussreich. Es gelang, Patienten mit aktiver TB von latent Infizierten und Nichtinfizierten mit hoher Sensitivität und Spezifität zu unterscheiden. Insgesamt wurden über 400 Stoffwechselprodukte untersucht, von denen etwa 20 Metabolite für eine robuste Diskriminierung der Populationen benötigt wurden. Hierzu gehörten insbesondere Mitglieder der Gruppen:
- Fettsäuren vom mittlerer Länge;
- Fibrinopeptide;
- Lysophosphatidylcholine;
- Kynurenine.
Die Definition einer Biosignatur, die das Risiko eines Krankheitsausbruchs in latent infizierten Gesunden voraussagen kann, steht noch aus. Hierzu wurden aber die wesentlichen Weichen gestellt. Mit finanzieller Unterstützung der Bill und Melinda Gates Stiftung wird von uns eine multizentrische retrospektive Studie in Afrika koordiniert. Im Zentrum des Interesses stehen Haushaltskontakte frisch diagnostizierter TB-Patienten. Die Rekrutierung der Haushaltskontakte ist bereits abgeschlossen. Etwa 4.500 Haushaltskontakte von rund 850 neu diagnostizierten Patienten mit aktiver Lungen-TB wurden in diese Studie aufgenommen. Direkt nach Aufnahme in die Studie sowie 6, 18 und 24 Monate danach wird diesen Probanden Blut abgenommen, die ab Mitte 2012 auf Biosignaturen hin untersucht werden. Zum Ende des Jahres 2010 wurden unter den Haushaltskontakten rund 70 Fälle mit aktiver TB diagnostiziert. Wir können somit davon ausgehen, dass etwa 3% der Studienteilnehmer in Verlauf der zwei Jahre dauernden Beobachtung eine aktive TB entwickeln werden, während 97% latent infiziert bleiben, also nicht erkranken. Nach Abschluss der Studie werden zwischen 90 und 115 TB-Fälle erwartet, je nachdem, wie viele Kontaktpersonen in der Studie bleiben. Diese Zahl reicht für eine robuste statistische Auswertung der Biosignaturen aus.