Forschungsbericht 2016 - Max-Planck-Institut für Kolloid- und Grenzflächenforschung
Wechselwirkung von Nanopartikeln mit Membranen und Vesikeln
Nanopartikel werden in der Medizin und Pharmakologie bereits heute als Wirkstoffträger, Diagnostika, Kontrastmittel und Implantate eingesetzt. Zwei besonders viel versprechende Einsatzgebiete in der Nano-Medizin sind Transportsysteme, die der zielgerichteten Verteilung von Wirkstoffen in bestimmten Geweben des menschlichen Körpers dienen, sowie magnetische Nanopartikel, die in der Krebstherapie zur Anwendung kommen, um durch Erwärmung Tumorzellen zu zerstören.
Versucht ein Nanopartikel beispielsweise als Wirkstoffträger in eine Zelle einzudringen, muss es zunächst die äußere Zellmembran überwinden. Dieser Prozess, auch Endozytose genannt, beginnt mit der Bindung des Nanopartikels an die Zellmembran. Danach muss die Membran über die Partikeloberfläche spreiten, bis sie das Partikel vollständig umhüllt. Ursache für diese Umhüllung des Partikels („particle engulfment“) sind die attraktiven Kräfte zwischen Membran und Partikel. Sie zwingen die Membran, sich lokal an die Partikelform anzupassen. Deshalb hängt dieser Prozess auch stark von den krümmungselastischen Eigenschaften der Membran ab.
Biologische und biomimetische Membranen bestehen aus molekularen Doppelschichten, in denen sich Lipide und Membranproteine so anordnen, dass die wasserlöslichen Molekülsegmente die Kontaktflächen zum Wasser bilden. Um wasserunlösliche Kanten zu vermeiden, schließen sich diese Doppelschichten zu blasenartigen Flächen, sogenannten Vesikeln, die zwei getrennte wässrige Kompartimente erzeugen. Ein besonders einfacher Fall liegt vor, wenn einerseits die beiden Monoschichten oder Blättchen der Doppelschicht aus der gleichen Lipid-Protein-Mischung bestehen und wenn andererseits die beiden wässrigen Kompartimente die gleiche ionische und molekulare Zusammensetzung besitzen. Dann ist die Doppelschicht symmetrisch und besitzt keine Vorzugskrümmung („spontaneous curvature“).Wenn Nanopartikel mit einer symmetrischen und gleichzeitig planaren Membran in Kontakt kommen, werden sie oberhalb einer bestimmten kritischen Größe vollständig von der Membran umhüllt. Kleinere Partikel hingegen haften trotz der anziehenden Kräfte überhaupt nicht an der Membran [1].
Biologische Membranen sind allerdings weder planar noch symmetrisch. Vielmehr besitzen sie fast immer eine komplexe, nicht sphärische Form mit variabler Krümmung. Zudem sind alle biologischen Membranen asymmetrisch, weil ihre inneren und äußeren Blättchen aus unterschiedlichen Lipidmischungen bestehen und mit verschiedenen im Wasser gelösten Stoffen in Kontakt kommen. Wie sich vor kurzem anhand von mathematischen Modellen herausstellte, beeinflussen sowohl die lokale Krümmung als auch die Asymmetrie der Membran die Umhüllung von Nanopartikeln maßgeblich [2-5].
Membran-Umhüllung eines Nanopartikels
 im Kontakt mit einer asymmetrischen Doppelschicht-Membran (rote und blaue Monoschicht) kann einen freien (F), partiell umhüllten (P) oder komplett umhüllten (C) Zustand erreichen. Bei letzterem bildet die Membran einen engen Hals zwischen den partikelgebundenen und den ungebundenen Membransegmenten.")
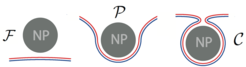
Abb.1: Ein kugelförmiges Nanopartikel (NP, grau) im Kontakt mit einer asymmetrischen Doppelschicht-Membran (rote und blaue Monoschicht) kann einen freien (F), partiell umhüllten (P) oder komplett umhüllten (C) Zustand erreichen. Bei letzterem bildet die Membran einen engen Hals zwischen den partikelgebundenen und den ungebundenen Membransegmenten.
Je nach der Bedeckung der Partikeloberfläche durch die Membran lassen sich drei Zustände eines Nanopartikels unterscheiden (siehe Abb. 1): Zustand F: Die Membran spreitet gar nicht über die Partikeloberfläche; Zustand P: partielle Umhüllung des Partikels durch die Membran; Zustand C: komplette Umhüllung des Partikels durch die Membran. Im Zustand C ist das partikelgebundene Membransegment mit der restlichen Membran durch einen engen Membranhals verbunden. Eine Kombination von analytischer Theorie und numerischen Rechnungen führt auf zwei universelle und relativ einfache Bedingungen, die angeben, für welchen Partikelgrößen die freien und die komplett umhüllten Zustände F und C stabil sind [2].
In Abhängigkeit von der Membranasymmetrie und der lokalen Membrankrümmung ergeben sich aus diesen beiden Stabilitätsbedingungen zwei kritische Partikelradien, R1 und R2. Kleine Partikel mit einem Radius kleiner als R1 sind frei und bleiben nicht an der Membran haften. Große Partikel mit einem Radius größer als R2 sind komplett umhüllt. Partikel mit Radien zwischen R1 und R2 sind entweder partiell umhüllt oder gehören zu einem bistabilen Bereich, in dem der freie und der komplett umhüllte Zustand gleichzeitig (meta)stabil sind. Demnach ergeben sich aus den beiden Stabilitätsbedingungen vier verschiedene Umhüllungsbereiche für ein einzelnes Nanopartikel.
Umhüllungsmuster von vielen Nanopartikeln
Die Stabilitätsbedingungen, die die Umhüllungsbereiche definieren, hängen von der lokalen Krümmung der Membran ab. Deshalb können verschiede Segmente einer nicht sphärischen Vesikelmembran zu verschiedenen Umhüllungsbereichen gehören. Die verschiedenen Segmente einer gegebenen Vesikelform definieren ein Umhüllungsmuster, das sich ausbildet, wenn die Vesikel mit vielen Nanopartikeln wechselwirkt. Insgesamt gibt es zehn unterschiedliche Muster, die jeweils aus bis zu drei verschiedenen Membransegmenten bestehen [3]. Auf einer zigarrenförmigen Vesikel können sich acht verschiedene Muster bilden, wenn man die Membran-Asymmetrie, das Haftvermögen und/oder die Partikelgröße variiert (siehe Abb. 2).
Krümmungsinduzierte Kräfte
 auf einer zigarrenförmigen Vesikel. Die Vorzugskrümmung der Membran ist positiv in (a) und negativ in (b). Die Muster bestehen aus vier verschiedenartigen Membransegmenten, die den vier möglichen Umhüllungszuständen der Partikel entsprechen: Einerseits (i) freie Segmente ohne haftende Partikel (rot) und (ii) bistabile Segmente (orange), auf denen freie und komplett umhüllte Partikelzustände koexistieren können, sowie andererseits (iii) Segmente mit partiell umhüllten (blau) und (iv) komplett umhüllten (grün) Partikeln. Die Änderung der Partikelgröße oder des Haftvermögens führt zu kontinuierlichen morphologischen Transformationen zwischen diesen Mustern.")
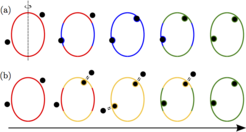
Abb.2: Umhüllungsmuster von Nanopartikeln (schwarz) auf einer zigarrenförmigen Vesikel. Die Vorzugskrümmung der Membran ist positiv in (a) und negativ in (b). Die Muster bestehen aus vier verschiedenartigen Membransegmenten, die den vier möglichen Umhüllungszuständen der Partikel entsprechen: Einerseits (i) freie Segmente ohne haftende Partikel (rot) und (ii) bistabile Segmente (orange), auf denen freie und komplett umhüllte Partikelzustände koexistieren können, sowie andererseits (iii) Segmente mit partiell umhüllten (blau) und (iv) komplett umhüllten (grün) Partikeln. Die Änderung der Partikelgröße oder des Haftvermögens führt zu kontinuierlichen morphologischen Transformationen zwischen diesen Mustern.
Typischerweise ist das Nanopartikel klein im Vergleich zur Vesikelgröße. Für diesen Fall wurde in der Abteilung `Theorie und Bio-Systeme’ am Max-Planck-Institut für Kolloid- und Grenzflächenforschung eine analytische Theorie entwickelt, die im Detail beschreibt, wie die Energielandschaft der Membran-Partikel-Systeme von der Vesikelgeometrie und den Materialparametern abhängt [4].
Die Theorie sagt insbesondere voraus, dass die Energie einer partiell umhüllten Partikel von der lokalen Krümmung der Vesikelmembran abhängt. Wenn die Vesikel die Form einer Kugel hat, dann ist die Membrankrümmung für alle Segmente der Vesikelmembran konstant und der Partikel bewegt sich in einer flachen Energielandschaft. Für alle anderen Vesikelformen hängt die Membrankrümmung dagegen von der Position des Membransegments ab. Deshalb führt die nicht sphärische Form einer Vesikelmembran auf Energiegradienten und damit auf krümmungsinduzierte Kräfte, die die Membran auf partiell umhüllte Nanopartikel ausübt. Wenn die Partikel von außen an die Vesikelmembran binden (Endozytose), treiben diese Kräfte die Partikel zu den am schwächsten gekrümmten Membransegmenten. Binden die Partikel dagegen von innen an die Membran, so wirken die Kräfte in Richtung zunehmender Membrankrümmung.
Die partielle Membranumhüllung eines Nanopartikels mit einer chemisch einheitlichen Oberfläche ist nur möglich, wenn die Partikelgröße und das Haftvermögen genau auf die anderen Membraneigenschaften abgestimmt sind. Im Gegensatz dazu werden sogenannte Janus-Partikel, die eine stark haftende und eine nicht haftende Oberflächenregion besitzen, immer partiell umhüllt. Deshalb lassen sich krümmungsinduzierte Kräfte am einfachsten für solche Janus-Partikel experimentell untersuchen (siehe Abb. 3).
 Zigarrenförmige Vesikel (grün-blau) in Kontakt mit chemisch homogenen Nanopartikeln (grau). In diesem Beispiel werden die Partikel an den stark gekrümmten Polen (grün) komplett, an den schwächer gekrümmten äquatorialen Regionen (blau) dagegen nur partiell umhüllt. In dem hier gezeigten endozytischen Fall sind die partiell umhüllten Partikel krümmungsinduzierten Kräften ausgesetzt, die in Richtung abnehmender Membrankrümmung wirken. Komplett umhüllte Partikel werden dagegen von diesen Kräften nicht beeinflusst. (b) Die gleiche Vesikel führt bei Januspartikeln mit einer stark haftenden (grau) und einer nicht haftenden (rot) Oberflächenregion überall auf der Vesikel zur partiellen Umhüllung. Aus diesem Grund werden diese Partikel immer durch krümmungsinduzierte Kräfte in Richtung Äquator getrieben.")
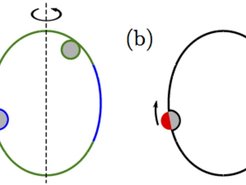
Abb. 3: (a) Zigarrenförmige Vesikel (grün-blau) in Kontakt mit chemisch homogenen Nanopartikeln (grau). In diesem Beispiel werden die Partikel an den stark gekrümmten Polen (grün) komplett, an den schwächer gekrümmten äquatorialen Regionen (blau) dagegen nur partiell umhüllt. In dem hier gezeigten endozytischen Fall sind die partiell umhüllten Partikel krümmungsinduzierten Kräften ausgesetzt, die in Richtung abnehmender Membrankrümmung wirken. Komplett umhüllte Partikel werden dagegen von diesen Kräften nicht beeinflusst. (b) Die gleiche Vesikel führt bei Januspartikeln mit einer stark haftenden (grau) und einer nicht haftenden (rot) Oberflächenregion überall auf der Vesikel zur partiellen Umhüllung. Aus diesem Grund werden diese Partikel immer durch krümmungsinduzierte Kräfte in Richtung Äquator getrieben.
Rezeptor-vermittelte Endozytose
Bislang wurden nur Membranen mit einer lateral homogenen molekularen Zusammensetzung betrachtet. Um die komplexeren Prozesse der Endozytose in echten Zellen nachzuahmen, muss man auch die haftinduzierte Entmischung von Membranmolekülen berücksichtigen. Diese Entmischung entsteht durch die unterschiedlichen Wechselwirkungen zwischen den verschiedenen Membrankomponenten und dem Nanopartikel und führt auf partikelgebundene und ungebundene Membransegmente, die sich in ihrer Biegesteifigkeit und Vorzugskrümmung unterscheiden [2]. Auf diese Weise konnten die experimentellen Daten für die Clathrin-vermittelte Endozytose von Goldpartikeln durch HeLa-Zellen erklärt werden. Die Daten zeigen, dass die Partikelaufnahme durch die Membran nicht-monoton von der Teilchengröße abhängt. Bei einem Durchmesser von 50 Nanometern, der der Vorzugskrümmung des Clathrin-bedeckten Membransegments entspricht, ist sie am größten.
Stabilisierung von engen Membranhälsen
Wie bereits erwähnt, führt ein komplett eingehülltes Partikel immer zu einem engen Membranhals (siehe Abb. 1). Derartige Hälse treten auch bei vielen anderen Membranprozessen auf. Wichtige Beispiele dafür sind die Knospung („budding“) und die Röhrenbildung von Festkörper-unterstützten Lipiddoppelschichten, die Bildung von extrazellulären und äußeren Membranvesikeln durch eukaryotische und prokaryotische Zellen, die Zytokinese während der Zellteilung oder die kollektive Aufnahme von vielen Partikeln in halskettenartigen Membranröhren (siehe Abb. 4). Ferner unterstützen in Zellen lokalisierte Kompressionskräfte oft die Bildung von engen Membranhälsen, indem sie direkt auf diese einwirken. Dies geschieht in der Regel mithilfe von Proteinen wie Dynamin bei der Endozytose oder Actomyosin bei der Zytokinese. Die Stabilität der Membranhälse lässt sich in all diesen Systemen durch eine vereinheitlichte Stabilitätsbedingung beschreiben, die vom Haftvermögen der Partikel, den Krümmungs-elastischen Eigenschaften der Membran, der lokalen Membrankrümmung sowie der Stärke der lokalisierten Kompressionskräfte abhängt [5].
 oder Kompressionskräften enge Membranhälse ausgebildet werden. (a) Knospung oder Röhrenbildung einer Festkörper-unterstützten Doppelschicht; (b) Bildung von großen Vesikeln aus der äußeren Zellmembran, die vom Aktinkortex unterstützt wird; (c) Umhüllung von vielen Nanopartikeln in eine halskettenähnliche Membranröhre; und (d) Stabilisierung eines Membranhalses durch einen kontraktilen Protenring (grün), der lokalisierte Kompressionskräfte auf den Hals ausübt.")
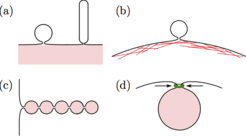
Abb. 4: Verschiedene Systeme, in denen auf Grund von haftfähigen Oberflächen (pink) oder Kompressionskräften enge Membranhälse ausgebildet werden. (a) Knospung oder Röhrenbildung einer Festkörper-unterstützten Doppelschicht; (b) Bildung von großen Vesikeln aus der äußeren Zellmembran, die vom Aktinkortex unterstützt wird; (c) Umhüllung von vielen Nanopartikeln in eine halskettenähnliche Membranröhre; und (d) Stabilisierung eines Membranhalses durch einen kontraktilen Protenring (grün), der lokalisierte Kompressionskräfte auf den Hals ausübt.
Unsere Arbeiten zeigen, dass die Endozytose von Nanopartikeln sehr empfindlich von der Vorzugskrümmung der Doppelschichtmembranen abhängt und dass sich daraus eine komplexe Abhängigkeit der Endozytose von der Teilchengröße ergibt. Die theoretischen Ergebnisse erklären die experimentell beobachtete Abhängigkeit für die Clathrin-vermittelte Endozytose von Goldpartikeln durch HeLa-Zellen. In diesem Fall ist die Vorzugskrümmung der Membran durch die Größe des Membran-verankerten Proteingerüsts bestimmt und führt auf eine maximale Partikelaufnahme für eine Teilchengröße von etwa 50 Nanometer. Da der zu Grunde liegende Mechanismus sehr allgemein ist, sollten sich auch für andere Formen der Endozytose optimale Teilchengrößen für die Partikelaufnahme ergeben. Wenn die Nanopartikel als Wirkstoffträger eingesetzt werden, kann man deren Wirksamkeit durch eine entsprechende Wahl der Teilchengröße optimieren.
Auch unsere vereinheitlichte Stabilitätsbedingung für Membranhälse stellt eine sehr allgemeine Beziehung zwischen experimentell beobachtbaren Größen dar. Diese Stabilitätsbedingung gilt insbesondere für die symmetrische Knospung von Zellmembranen, wie sie bei der Zellteilung auftritt. Hier ergeben sich interessante Anwendungen im Rahmen der Synthetischen Biologie.