Forschungsbericht 2016 - Max-Planck-Institut für Hirnforschung
Computerbasierte Analyse und Modellierung von umfangreichen aus dem Gehirn gewonnenen Daten
Abteilung Computation in Neural Circuits
Abteilung Memory and Navigation Circuits
Die Plastizität neuraler Schaltkreise
Neurale Schaltkreise stehen einem fundamentalen Problem gegenüber: Wie kann Erfahrung im Rahmen der Plastizität zu Veränderung und Verbesserung der Verschaltungen in Netzwerken führen, ohne dass gleichzeitig die Funktionsstabilität verloren geht? Diese beiden Anforderungen werden von zwei verschiedenen Formen synaptischer Plastizität erfüllt: Hebb’sche Mechanismen, die synaptische Konnektivität Synapsen-spezifisch verändern, und homöostatische Mechanismen, die eine stabile Funktion aufrechterhalten, indem die allgemeinen synaptischen Gewichte und das Gleichgewicht zwischen Exzitation und Inhibition kontinuierlich global angepasst werden [1]. Wie homöostatische Mechanismen innerhalb komplexer Mikroverschaltungen interagieren, um deren Funktion aufrechtzuerhalten, ist immer noch weitgehend unbekannt.
Dieser Frage gehen die Max-Planck-Forscher im Rahmen einer internationalen Zusammenarbeit nach. Dazu verwenden sie die theoretische und experimentbasierte Charakterisierung kortikaler synaptischer Plastizität und Netzwerkdynamiken während sensorischer Deprivation im visuellen Kortex von Nagetieren. Das von uns verwendete Modellsystem ist der visuelle Kortex (V1) von Nagetieren (Ratten und Mäusen) während der klassischen „kritischen Phase“ des visuellen Systems. Dieser etwa vom postnatalen Tag P22 – P33 dauernde Zeitraum zeichnet sich durch einen hohen Grad von Plastizität aus, die durch die Manipulation visueller Erfahrung eingeleitet werden kann (Abb. 1).
![Abb. 1: Zeitachse der Induktion der MD (Monokulare Deprivation) und der chronischen Aufzeichnungen (oben). P15: Augenöffnung, P21: Implantat, ab P24: Aufzeichnung, ab P27: Lidverschluss. Während die Feuerraten der exzitatorischen "Regulär feuernden Zellen" (RSUs, vorwiegend Pyramidenzellen) in Schicht 4 zwei Tage nach monokularer Deprivation abfallen, sinken die Feuerraten inhibitorischer Neurone - sowohl der RSUs als auch mutmaßlicher "schnell feuernder Zellen" (FS) in allen Schichten bereits einen Tag zuvor. (Abb. verändert nach [2]).](/11636196/original-1508158529.jpg?t=eyJ3aWR0aCI6MzQxLCJmaWxlX2V4dGVuc2lvbiI6ImpwZyIsIm9ial9pZCI6MTE2MzYxOTZ9--68441974a65af03472fed8bb8811e09a5cbba177 "Abb. 1: Zeitachse der Induktion der MD (Monokulare Deprivation) und der chronischen Aufzeichnungen (oben). P15: Augenöffnung, P21: Implantat, ab P24: Aufzeichnung, ab P27: Lidverschluss. Während die Feuerraten der exzitatorischen \"Regulär feuernden Zellen\" (RSUs, vorwiegend Pyramidenzellen) in Schicht 4 zwei Tage nach monokularer Deprivation abfallen, sinken die Feuerraten inhibitorischer Neurone - sowohl der RSUs als auch mutmaßlicher \"schnell feuernder Zellen\" (FS) in allen Schichten bereits einen Tag zuvor. (Abb. verändert nach [2]).")
![Abb. 1: Zeitachse der Induktion der MD (Monokulare Deprivation) und der chronischen Aufzeichnungen (oben). P15: Augenöffnung, P21: Implantat, ab P24: Aufzeichnung, ab P27: Lidverschluss. Während die Feuerraten der exzitatorischen "Regulär feuernden Zellen" (RSUs, vorwiegend Pyramidenzellen) in Schicht 4 zwei Tage nach monokularer Deprivation abfallen, sinken die Feuerraten inhibitorischer Neurone - sowohl der RSUs als auch mutmaßlicher "schnell feuernder Zellen" (FS) in allen Schichten bereits einen Tag zuvor. (Abb. verändert nach [2]).](/11636196/original-1508158529.jpg?t=eyJ3aWR0aCI6MjQ2LCJvYmpfaWQiOjExNjM2MTk2fQ%3D%3D--8a5dd1653b5ef0aa99c61edb0a621a7e7685a8c5)
Abb. 1: Zeitachse der Induktion der MD (Monokulare Deprivation) und der chronischen Aufzeichnungen (oben). P15: Augenöffnung, P21: Implantat, ab P24: Aufzeichnung, ab P27: Lidverschluss. Während die Feuerraten der exzitatorischen "Regulär feuernden Zellen" (RSUs, vorwiegend Pyramidenzellen) in Schicht 4 zwei Tage nach monokularer Deprivation abfallen, sinken die Feuerraten inhibitorischer Neurone - sowohl der RSUs als auch mutmaßlicher "schnell feuernder Zellen" (FS) in allen Schichten bereits einen Tag zuvor. (Abb. verändert nach [2]).
Unser Ziel ist es, herauszufinden, in welchen Zeiträumen und über welche Mechanismen die synaptische und homöostatische Plastizität in diesen visuellen Mikroschaltkreisen als Folge von Aktivitätsmanipulation wirken, sowie die Veränderungen der Netzwerkdynamiken und Funktion zu beschreiben, die von ihnen eingeleitet werden. Ausgehend von detaillierten synaptischen Messungen der Exzitation und Inhibition, die durch afferente Eingänge (vom Thalamus kommend) und intrinsische Verbindungen (innerhalb der kortikalen Verbindungen) ausgelöst werden, beides sowohl unter normalen als auch unter deprivierten Bedingungen, werden Modelle des kortikalen Netzwerks entwickelt, um die resultierenden Netzwerk-Dynamiken zu erforschen, die aus den Veränderungen von afferenten und intrinsischen Verschaltungen nach der Deprivation hervorgehen.
Außerdem untersuchen wir die Natur der Plastizitätsmechanismen, die die kortikalen Kreisläufe neu strukturieren, wobei wir uns auf zwei Zeiträume konzentrieren: 1 bis 2 Tage nach der Deprivation sowie 6 Tage nach der Deprivation. Zusätzlich analysieren wir die experimentell gemessene Aktivität, die von vielen gleichzeitig untersuchten Zellen im deprivierten Kortex erhoben wurde. Dadurch können wir festlegen, welche Eigenschaften (z. B. Feuerraten, Kreuzkorrelationen u. a.) die Netzwerkdynamiken unserer Modelle aufweisen sollten. Mit unseren Modellen bestimmen wir die notwendigen synaptischen Veränderungen, welche die experimentell gemessenen Veränderungen der Netzwerkeigenschaften abbilden: zum Beispiel gehen die Feuerraten von inhibitorischen Neuronen einen Tag nach der Deprivation zurück und erholen sich am folgenden Tag, wenn die exzitatorischen Feuerraten zurückgehen, die sich dann innerhalb von 6 Tagen, also einen längeren Zeitraum, ebenfalls erholen (Abb. 1, [2]). Die Ergebnisse der Modellierung erlauben Prognosen für die Wirkungsweise des Netzwerks, in dem spezifische exzitatorische und inhibitorische synaptische Veränderungen zu den gemessenen Netzwerkeigenschaften beitragen. Die Kooperationspartner testen diese Voraussagen dann in realen biologischen Netzwerken und stellen zusätzliche Vorgaben bereit, um die Modelle zu verfeinern.
Parallele Pfade der visuellen Verarbeitung in der Retina
Seit den Arbeiten des spanischen Mediziners Ramón y Cajal (1852-1934) wissen wir, dass sich über die verschiedenen Hirnregionen viele verschiedene Zelltypen verteilen. Einer der am besten charakterisierten neuronalen Schaltkreise, um die Vorteile dieser Vielfalt an Zellarten bei der Reizverarbeitung zu verstehen, ist die Retina. In den vergangenen Jahren haben verschiedene Beweislinien gezeigt, dass retinale Ganglienzellen (die Zellen, die visuelle Informationen vom Auge zum Gehirn leiten) eine große Diversität in Bezug auf ihre Größe und anatomische Struktur aufweisen [3]. Bei vielen dieser Typen handelt es sich nicht wie bisher angenommen nur um Lichtdetektoren, sondern sie reagieren auf verschiedene visuelle Eigenschaften, zum Beispiel Bewegung und Farbe [4]. Die an nachgeschaltete Hirnregionen gesendeten Informationen sind daher parallelisiert und bereits maßgeblich nichtlinear vorverarbeitet. Um die Diversität der Zelltypen zu verstehen, muss man daher über herkömmliche Ansätze und lineare Modelle hinausgehen. Unter Berücksichtigung neuer experimenteller Ergebnisse verwenden die Forscher theoretische Ansätze aus der Informationstheorie, um neue Einblicke in die frühe visuelle Verarbeitung zu erlangen.
 wird von einer ON-Zelle (oben, Mitte) und einer OFF-Zelle (oben rechts) kodiert. Dabei bezeichnet ON die Reaktion auf positive Helligkeitsänderungen, OFF die Reaktion auf negative Helligkeitsänderungen. Die jeweiligen Informationen für eine Zellpopulation lassen sich als Funktion der beschränkten maximalen Feuerrate der Zellen darstellen (unten links). Die jeweiligen Informationen sind identisch für jede Kombination aus ON und OFF-Zellen und hängen nicht von der Stimulusverteilung ab (unten rechts). Die Verwendung einer alternativen Maßeinheit der Verschlüsselungseffizienz, der mittlere quadratische Fehler der optimalen linearen Auslesung, erzeugt eine starke Abhängigkeit der optimalen Anzahl an ON- und OFF-Zellen von den Asymmetrien der Stimulusverteilung P(s), angezeigt durch die unterschiedlichen Farben der Kurven.")
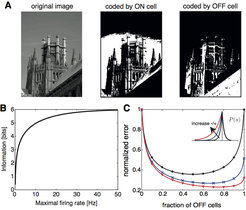
Eine bekannte Hypothese für die Informationsverarbeitung in peripheren sensorischen Systemen wie der Retina ist die Hypothese der effizienten Kodierung, die besagt, dass neurale Systeme dafür optimiert sind, ein Maximum an Informationen über sensorische Reize zu übertragen, das durch biologische Einschränkungen (z.B. biophysikalischer und metabolischer Art) begrenzt ist. Diese Hypothese wird verwendet, um die möglichen Vorteile von Zellartendiversität in der Retina zu erklären. Mit diesem Ansatz konnte das Vorhandensein zweier Pfade für die Verarbeitung gegensätzlicher Kontrastsignale erklärt werden, nämlich den ON- (Reaktion auf positive Helligkeitsänderungen) und OFF-Antworten (Reaktion auf negative Helligkeitsänderungen) (Abb. 2, [5]).
Computerbasierte Modelle sind ein leistungsfähiges, die Experimente ergänzendes Werkzeug; die meisten Modelle sind jedoch bisher von der Kodierung statischer Stimuli, die sich zeitlich nicht ändern, ausgegangen. Aus der Integration dynamischer Eigenschaften verhaltensrelevanter natürlicher Stimuli (zum Beispiel sich bewegender Objekte) lassen sich Prognosen darüber ableiten, wie retinale Zellen beschaffen sein müssen, um für diese Stimuli optimal zu kodieren. Dabei konzentrieren sich die Forscher auf zwei Klassen retinaler Zellen: retinale Ganglienzellen und bipolare Zellen. Ziel der Arbeit ist es, herauszufinden, inwieweit die Komplexität natürlicher Stimuli das Auftreten diverser Berechnungen in der Retina beeinflusst.
Entschlüsselung der neuralen Codes für räumliche Navigation
Wir leben in einem geometrischen Raum. Unser alltägliches Leben hängt stark von der Fähigkeit zu räumlicher Navigation ab. Wenn wir zum Beispiel planen, zu einem bestimmten Ort zu gelangen, müssen wir eine geeignete Route abschätzen, die von unserer aktuellen Position bis zum Ziel führt. Erstaunlicherweise können wir dies quasi sofort und ohne große Anstrengungen bewerkstelligen. Da sich ein Zielort typischerweise außerhalb des durch die Sinne wahrnehmbaren Bereiches befindet, dürfte die Routenplanung im Gehirn ohne direkte Leitung durch Sinnessignale implementiert werden. Jahrzehnte der neurowissenschaftlichen Forschung haben Neuronen identifiziert, die die Position oder Kopfausrichtung eines Tieres repräsentieren, und so ein internes Kartensystem im Gehirn konstituieren [6]. Es ist jedoch noch immer weitgehend unklar, wie das Gehirn diese Informationen zur Aktionsplanung zum Erreichen eines Zielorts verwenden kann. Die Frankfurter Max-Planck-Forschergruppe beschäftigt sich mit dieser Frage, indem sie sehr dichte elektrische Nervenzellableitungen in sich normal verhaltenden Ratten (Abb. 3) in Kombination mit optogenetischen Manipulationen neuronaler Schaltkreise verwendet.
 überwacht, während das Tier in einer Verhaltensbox (behavior box) eine räumliche Navigationsaufgabe ausführt. Die Bilder rechts zeigen ein Beispiel des Vorgehens bei der Datenanalyse. Aus Elektroden im Gehirn werden Multikanal-Spannungsspuren (multichannel voltage traces) gewonnen, aus denen sich Spike-Aktivität von einzelnen Neuronen extrahieren lassen. Diese ermöglichen es, Korrelationen neuraler Aktivität bei einem bestimmten Aspekt von Verhaltensweisen zu untersuchen. Die Aufzeichnungen rechts unten zeigen eine Beispielanalyse von Neuronen im Hippocampus, die an einen bestimmten Ort in ihrer Umgebung feuern, sogenannte „Ortszellen“. Die blauen Spuren (links) sind die Bewegungsbahnen des Tieres; die roten Punkte darin zeigen die Spike-Positionen an. Rechts: Eine farbkodierte Spike-Landkarte desselben Neurons, das links zu sehen ist.")
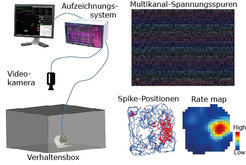
Abb. 3: Schema der experimentellen Methoden. Das linke Bild zeigt einen allgemeinen Experimentaufbau: Das Verhalten und die neurale Aktivität des Tieres werden durch eine Videokamera und ein Aufzeichnungssystem (recording system) überwacht, während das Tier in einer Verhaltensbox (behavior box) eine räumliche Navigationsaufgabe ausführt. Die Bilder rechts zeigen ein Beispiel des Vorgehens bei der Datenanalyse. Aus Elektroden im Gehirn werden Multikanal-Spannungsspuren (multichannel voltage traces) gewonnen, aus denen sich Spike-Aktivität von einzelnen Neuronen extrahieren lassen. Diese ermöglichen es, Korrelationen neuraler Aktivität bei einem bestimmten Aspekt von Verhaltensweisen zu untersuchen. Die Aufzeichnungen rechts unten zeigen eine Beispielanalyse von Neuronen im Hippocampus, die an einen bestimmten Ort in ihrer Umgebung feuern, sogenannte „Ortszellen“. Die blauen Spuren (links) sind die Bewegungsbahnen des Tieres; die roten Punkte darin zeigen die Spike-Positionen an. Rechts: Eine farbkodierte Spike-Landkarte desselben Neurons, das links zu sehen ist.
Das Hauptaugenmerk liegt dabei auf den funktionalen Interaktionen zwischen zwei Hirnregionen: dem präfrontalen Kortex, der dafür bekannt ist, für die Planung von Handlungen wichtig zu sein, und dem Hippocampus, der wichtig für das Gedächtnis und die räumliche Kodierung ist (Abb. 4, [7]). Ziel der Arbeiten ist es, herauszufinden, wie diese neuralen Schaltkreise verwendet werden, um so die Beziehung zwischen der geplanten Bewegung des Tieres und dem Zielort zu erforschen. Die Studie wird nicht nur Aufschluss über Navigations-Schaltkreise geben, sondern auch allgemeine Einblicke bezüglich der Frage gewähren, wie das Gehirn gespeicherte Informationen in den Schaltkreisen für Verhaltensplanung verwendet.
Die Rolle subkortikaler Afferenzen für das verhaltensabhängige funktionales Verkoppeln von Hirnregionen
Parallele Berechnung gilt als operationelles Schlüsselmerkmal des Gehirns, um komplexe Berechnungen schnell und effizient durchzuführen. Es ist bekannt, dass jede Hirnregion gewissermaßen eine spezialisierte Rolle ausübt: der Hippocampus spielt eine Schlüsselrolle beim Verarbeiten und Abrufen von Informationen; der präfrontale Kortex ist dafür bekannt, dass er für Handlungsplanungen und Entscheidungen wichtig ist. Damit ein Tier eine bestimmte verhaltensbezogene Aufgabe ausführen kann, ist ein Zusammenwirken der Hirnstrukturen wahrscheinlich essentiell. Wie kann der Hippocampus zum Beispiel eine angemessene Handlung implementieren, um die verhaltensbezogenen Entscheidungen im präfrontalen Kortex zu unterstützen?

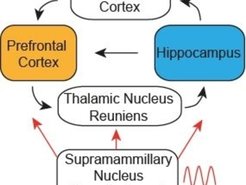
Die Forschergruppe geht diesen Fragen nach, indem sie die funktionale Interaktion zwischen dem Hippocampus und subkortikalen Strukturen untersucht. Wie erstmals von Papez (1937) angeregt, war der Hippocampus lange dafür bekannt, mit den Bereichen des Thalamus und des Hypothalamus in reziproker anatomischer Interaktion zu stehen [8], und viele klinische Fälle haben gezeigt, dass Schäden an diesem subkortikalen Bereich zu schweren Gedächtnisdefiziten führen, ein ähnliches Symptom wie bei Patienten mit Schädigung des Hippocampus [9]. Unser vorläufiges Ergebnis deutet darauf hin, dass der Nucleus supramammillaris im Hypothalamus eine entscheidende Rolle für die zeitliche Koordination von Aktionspotenzialen im Theta-Rhythmus im präfrontal-thalamo-hippocampalen Kreislauf spielt, der für das Verhalten bei der räumlichen Navigation wichtig ist (Abb. 4). Da die zeitliche Koordination einen starken Einfluss auf die Richtung des Signalflusses zwischen den Strukturen hat [10], können subkortikale Strukturen eine Schlüsselrolle in der dynamischen operationalen Regulierung des Hippocampus für Gedächtnisspeicherung und -abrufung spielen.